Comparison of Toxicity and Disposition of Cadmium Chloride and Cadmium- Metallothionein in Rats Promotoren: Dr
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(A) 1) at First a Pair of Each 10G. of Sample Was Extracted with Ether by the Soxhlet's Apparatus for a Week
52 [vol. 6, THE DISTRIBUTION OF PHOSPHORUS IN COW'S MILK AND THE SCHEME FOR THE SEPARATION OF PHOSPHATIDES By Rinjiro SASAKI (From the Institute of Agricultural Chemistry, College of Agriculture, Tokyo Imperial University) (Received 9th September 1930) A. The Distribution of Phosphorus in Cow's milk In order to extract the phosphatides, it is necessary first to determine the distribution of phosphorus in cow's milk. Liquid milk can not be extracted with ether or with other organic solvents owing to its large content of water. The substance applied in this experiment is the milk powder, dried by the Buflovak drum drier below 70•Ž. Its content of moisture is 4.498%, deter mined by the method of drying in the air bath of 105•Ž. The content of total phosphorus was determined(5) in the ash of sample which was burned with a little of fusing mixture. In 100g. of milk powder In 100g. of dry matter Total phosphorus 0.745g. 0.781g. The amounts of phosphorus, which are soluble in the various organic sol vents, were determined by extracting with three solvents in the following orders. (A) Ether-Acetone-Alcohol (B) Acetone-Ether-Alcohol (C) Alcohol-Ether-Acetone Experiment (A) 1) At first a pair of each 10g. of sample was extracted with ether by the Soxhlet's apparatus for a week. After the extraction had been comp leted, the extract was freed from ether and weighed. In 100g.of milkpowder In 100g.of dry matter Total ether matters soluble 3.609g. 3.779g. 2) The above extract was dissolved in a very little of ether. -

United States Patent Office Patented Apr
3,030,421 United States Patent Office Patented Apr. 17, 1962 2 3,030,421 perature and under a reduced pressure. In the case of PROCESS FOR PREPARNG TRHYDROXY most of the catalysts the reaction products, which are free METHYL-PHOSPHINE s from solvents, solidify already when being cooled to room Martin Reuter and Ludwig Ortane, Frankfurt an Main, temperature. If, in the case of some catalysts, they Germany, assignors to Farbwerke Hoechst Aktienger 5 solidify only at a lower temperature and still contain to sellschaft vormas Meister Lucius & Briining, Frank a greater extent oily by-products and/or phosphonium furt am Main, Germany, a corporation of Gerinally hydroxide, they can be separated from the latter by filter No Drawing. Fied Jaa. 14, 1958, Ser. No. 708,764 ing or pressing. - - - Claims priority, application Germany Jan. 23, 1957 It is to be assumed that the crystalline main product 6 Canas. (C. 260-606.5) O of the present invention constitutes the hitherto unknown We have found that a new and valuable phosphorus trihydroxymethyl-phosphine. Main and by-products are compound carrying hydroxymethyl groups at the phos easily soluble in water and methanol and sparingly solu phorus atom can be prepared by reacting 1 mol of formal ble in fat dissolvers. dehyde with 4 mol of phosphine, preferably in the pres The reaction products of the invention can be used as ence of water, and in the presence of Small quantities of 5 insecticides, additives for lubricants, flame-proofing agents finely distributed metals that do not belong to the alkali for wood and textiles and as intermediates for these sub metals or alkaline earth metals and/or their compounds Staces. -

Thermodynamic Properties of Mixtures of Aqueous Solutions of Hydrochloric Acid and Cadmium Chloride, Copper Chloride, Manganese Chloride, and Zinc Chloride
Western Michigan University ScholarWorks at WMU Master's Theses Graduate College 12-1993 Thermodynamic Properties of Mixtures of Aqueous Solutions of Hydrochloric Acid and Cadmium Chloride, Copper Chloride, Manganese Chloride, and Zinc Chloride Samia A. Kosa Follow this and additional works at: https://scholarworks.wmich.edu/masters_theses Part of the Chemistry Commons Recommended Citation Kosa, Samia A., "Thermodynamic Properties of Mixtures of Aqueous Solutions of Hydrochloric Acid and Cadmium Chloride, Copper Chloride, Manganese Chloride, and Zinc Chloride" (1993). Master's Theses. 4363. https://scholarworks.wmich.edu/masters_theses/4363 This Masters Thesis-Open Access is brought to you for free and open access by the Graduate College at ScholarWorks at WMU. It has been accepted for inclusion in Master's Theses by an authorized administrator of ScholarWorks at WMU. For more information, please contact [email protected]. THERMODYNAMIC PROPERTIES OF MIXTURES OF AQUEOUS SOLUTIONS OF HYDROCHLORIC ACID AND CADMIUM CHLORIDE, COPPER CHLORIDE, MANGANESE CHLORIDE, AND ZINC CHLORIDE by Samia A. Kosa A Thesis Submitted to the Faculty of The Graduate College in partial fulfillment of the requirements for the Degree of Master of Arts Department of Chemistry Western Michigan University Kalamazoo, Michigan December 1993 ACKNOWLEDGEMENTS I wish to express my deep gratitude and sincere appreciation to my committee advisor, Dr. Donald Schriber, for his continual guidance, direction, and assistance in this research. I would also like to acknowledge the help of the other committee members, Dr. Thomass Houser and Dr. Ralph Steinhaus, towards this work. My family continuously provided me support and encouragement and to them I owe more than I can say. -

Download Author Version (PDF)
Journal of Materials Chemistry A Accepted Manuscript This is an Accepted Manuscript, which has been through the Royal Society of Chemistry peer review process and has been accepted for publication. Accepted Manuscripts are published online shortly after acceptance, before technical editing, formatting and proof reading. Using this free service, authors can make their results available to the community, in citable form, before we publish the edited article. We will replace this Accepted Manuscript with the edited and formatted Advance Article as soon as it is available. You can find more information about Accepted Manuscripts in the Information for Authors. Please note that technical editing may introduce minor changes to the text and/or graphics, which may alter content. The journal’s standard Terms & Conditions and the Ethical guidelines still apply. In no event shall the Royal Society of Chemistry be held responsible for any errors or omissions in this Accepted Manuscript or any consequences arising from the use of any information it contains. www.rsc.org/materialsA Page 1 of 9 Journal of Materials Chemistry A ARTICLE JMCA Safer Salts for CdTe Nanocrystal Solution Processed Solar Cells: The Dual Roles of Ligand Exchange and Grain Growth Received 00th January 20xx, a b c d e Accepted 00th January 20xx Troy K. Townsend, † William B. Heuer, Edward E. Foos, Eric Kowalski, Woojun Yoon and Joseph G. Tischler e DOI: 10.1039/x0xx00000x Inorganic CdSe/CdTe nanocrystals for solid-state photovoltaic devices are typically sintered into a bulk-like material after www.rsc.org/ annealing in the presence of solid cadmium chloride. -
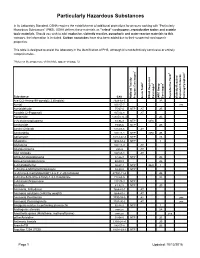
Particularly Hazardous Substances
Particularly Hazardous Substances In its Laboratory Standard, OSHA requires the establishment of additional protections for persons working with "Particularly Hazardous Substances" (PHS). OSHA defines these materials as "select" carcinogens, reproductive toxins and acutely toxic materials. Should you wish to add: explosive, violently reactive, pyrophoric and water-reactve materials to this category, the information is included. Carbon nanotubes have also been added due to their suspected carcinogenic properties. This table is designed to assist the laboratory in the identification of PHS, although it is not definitively conclusive or entirely comprehensive. *Notes on the proper use of this table appear on page 12. 1 6 5 2 3 4 Substance CAS National Toxicity National Program Carcinogen Toxin Acute Regulated OSHA Carcinogen Group IARC Carcinogen Toxin Reproductive Violently Reactive/ Explosive/Peroxide Forming/Pyrophoric A-a-C(2-Amino-9H-pyrido[2,3,b]indole) 2648-68-5 2B Acetal 105-57-7 yes Acetaldehyde 75-07-0 NTP AT 2B Acrolein (2-Propenal) 107-02-8 AT Acetamide 126850-14-4 2B 2-Acetylaminofluorene 53-96-3 NTP ORC Acrylamide 79-06-6 NTP 2B Acrylyl Chloride 814-68-6 AT Acrylonitrile 107-13-1 NTP ORC 2B Adriamycin 23214-92-8 NTP 2A Aflatoxins 1402-68-2 NTP 1 Allylamine 107-11-9 AT Alkylaluminums varies AT Allyl Chloride 107-05-1 AT ortho-Aminoazotoluene 97-56-3 NTP 2B para-aminoazobenzene 60-09-3 2B 4-Aminobiphenyl 92-67-1 NTP ORC 1 1-Amino-2-Methylanthraquinone 82-28-0 NTP (2-Amino-6-methyldipyrido[1,2-a:3’,2’-d]imidazole) 67730-11-4 2B -

The Effect of Sargassum Angustifolium Ethanol Extract on Cadmium Chloride-Induced Hypertension in Rat
Research Journal of Pharmacognosy (RJP) 8(1), 2021: 81-89 Received: 31 Oct 2020 Accepted: 17 Dec 2020 Published online: 19 Dec 2020 DOI: 10.22127/RJP.2020.255203.1637 Original article The Effect of Sargassum angustifolium Ethanol Extract on Cadmium Chloride-Induced Hypertension in Rat Leila Safaeian1* , Afsaneh Yegdaneh2, Masoud Mobasherian1 1Department of Pharmacology and Toxicology, Isfahan Pharmaceutical Sciences Research Center, School of Pharmacy and Pharmaceutical Sciences, Isfahan University of Medical Sciences, Isfahan, Iran. 2Department of Pharmacognosy, School of Pharmacy and Pharmaceutical Sciences, Isfahan University of Medical Sciences, Isfahan, Iran. Abstract Background and objectives: Sargassum angustifolium is a brown alga in southwestern coastline of Persian Gulf. Regarding the presence of various bioactive compounds and evidence of antihypertensive effects in other species of Sargassum, we evaluated the effect of S. angustifolium ethanol extract in CdCl2-induced hypertension in Wistar rats. Methods: Alga extract was prepared by maceration method using 70% ethanol and assessed for total phenolics and salt content. CdCl2 (1.5 mg/kg/day) was administered intraperitoneally to the rats for two weeks. Treatment groups received S. angustifolium extract (20, 40 and 80 mg/kg) or nifedipine (10 mg/kg) orally and simultaneously were given CdCl2 for two weeks. Systolic blood pressure (SBP) and heart rate were measured using tail-cuff method. Total antioxidant capacity, urea, creatinine, electrolytes including sodium, potassium, calcium and chloride were estimated in blood samples. The weight and histopathology of kidney tissues were also evaluated. Results: The content of total phenolic as gallic acid equivalent and the salt as NaCl was 67.42 ± 9.5 mg/g and 6.9 g/100 g in dried ethanol extract, respectively. -

Table of Contents
UNIVERSITY OF MISSOURI-KANSAS CITY CHEMICAL MANAGEMENT PLAN Revised May 2016 UMKC CHEMICAL MANAGEMENT PLAN This document constitutes the Chemical Management Plan (CMP) for the University of Missouri-Kansas City (UMKC). It was developed by the Environmental Health and Safety Department (EHS), to ensure the safe and proper use of hazardous and non- hazardous chemicals and to comply with applicable governmental regulations addressing the disposal of these chemicals. In addition, it was developed to foster waste minimization, and to provide the faculty and the staff with a management program to reduce the potential for accidents involving hazardous chemicals and/or wastes. Elements of the CMP include: a. a procedure for identifying potential or actual hazardous chemicals or wastes b. a procedure for periodic reexamination of those hazardous chemicals or wastes identified by the procedure in (a.) above as well as a systematic method for identification and evaluation of any new potential or actual hazardous chemicals or wastes c. procedures for labeling, and inventorying hazardous chemicals or wastes d. a procedure for identification and training of personnel directly responsible for ensuring that (a.), (b.), and (c.) are implemented e. a procedure for monitoring, recording, and reporting compliance with the CMP f. a procedure by which information generated by the CMP is provided to the persons performing waste analyses Each element is addressed as part of the complete CMP in the following paragraphs. 4 Table of Contents 1 Definitions 7 2 Identification -

NXP Semiconductors NXP List of Hazardous Substances in Products
NXP Semiconductors NX3-00119 NXP List of Hazardous Substances Sustainability 01-October-2012 in Products and Packaging Sustainability Page 1 of 7 Supersedes: NX3-00119 01-October-2010 Table of contents 1 Objectives ............................................................................................................................. 2 2 Scope ................................................................................................................................... 2 3 Responsibilities, Records & Risks......................................................................................... 2 3.1 Roles and Responsibilities .................................................................................................... 2 3.2 Records ................................................................................................................................ 2 3.3 Risks .................................................................................................................................... 3 4 References ........................................................................................................................... 3 5 Definitions/Abbreviations ...................................................................................................... 3 5.1 Definitions ............................................................................................................................. 3 5.2 Abbreviations ....................................................................................................................... -

The Reaction of Diphenyl Cadium with Phosphorus Trichloride
Union College Union | Digital Works Honors Theses Student Work 6-1940 The er action of diphenyl cadium with phosphorus trichloride Alfred Henry Case Union College - Schenectady, NY Follow this and additional works at: https://digitalworks.union.edu/theses Part of the Chemistry Commons Recommended Citation Case, Alfred Henry, "The er action of diphenyl cadium with phosphorus trichloride" (1940). Honors Theses. 1745. https://digitalworks.union.edu/theses/1745 This Open Access is brought to you for free and open access by the Student Work at Union | Digital Works. It has been accepted for inclusion in Honors Theses by an authorized administrator of Union | Digital Works. For more information, please contact [email protected]. The Reaction of Diphenyl Cadmium wj_th Phosphorus Trichloride J A thesis presented to the Department of Chem• istry of Union College, in partial fulfillment of the requirements for the Degree of Bachelor of Science in Chemistry, by ;,-. J ..,' ,., .... -. ' - " .. ' .. -..,, .. June, 1940 '"' ' - ' -1- Historical Any study of the stereochernistry of tricovalent atoms would necessarily involve members of the third and fifth periodic groups. Tertiary phosphines with heavy grol .pa would be most valuable for this work since phosphorus seems to be better suited to ezhi- bit asymmetry than any other element 01' the fifth group.<24) It is believed that the heavy groups would slow down the oscillations of the phosphorus atom and make possible the resolution of the optical isomers .. However, this work has been retarded by the lack of a convenient means of preparing asymmetrical sub- stituted triaryl phosphines. The usual method of preparing triaryl phosphines is by the reaction of the appropriate phosphenyl chloride with a Grignard reagent. -

SUMMARY of PARTICULARLY HAZARDOUS SUBSTANCES (By
SUMMARY OF PARTICULARLY HAZARDOUS SUBSTANCES (by alpha) Key: SC -- Select Carcinogens RT -- Reproductive Toxins AT -- Acute Toxins SA -- Readily Absorbed Through the Skin DHS -- Chemicals of Interest Revised: 11/2012 ________________________________________________________ ___________ _ _ _ _ _ _ _ _ _ _ _ ||| | | | CHEMICAL NAME CAS # |SC|RT| AT | SA |DHS| ________________________________________________________ ___________ | _ | _ | _ | _ | __ | | | | | | | 2,4,5-T 000093-76-5 | | x | | x | | ABRIN 001393-62-0 | | | x | | | ACETALDEHYDE 000075-07-0 | x | | | | | ACETAMIDE 000060-35-5 | x | | | | | ACETOHYDROXAMIC ACID 000546-88-3 ||x| | x | | ACETONE CYANOHYDRIN, STABILIZED 000075-86-5 | | | x | | x | ACETYLAMINOFLUORENE,2- 000053-96-3 | x | | | | | ACID MIST, STRONG INORGANIC 000000-00-0 | x | | | | | ACROLEIN 000107-02-8 | | x | x | x | | ACRYLAMIDE 000079-06-1 | x | x | | x | | ACRYLONITRILE 000107-13-1 | x | x | x | x | | ACTINOMYCIN D 000050-76-0 ||x| | x | | ADIPONITRILE 000111-69-3 | | | x | | | ADRIAMYCIN 023214-92-8 | x | | | | | AFLATOXIN B1 001162-65-8 | x | | | | | AFLATOXIN M1 006795-23-9 | x | | | | | AFLATOXINS 001402-68-2 | x | | x | | | ALL-TRANS RETINOIC ACID 000302-79-4 | | x | | x | | ALPRAZOMAN 028981-97-7 | | x | | x | | ALUMINUM PHOSPHIDE 020859-73-8 | | | x | | x | AMANTADINE HYDROCHLORIDE 000665-66-7 | | x | | x | | AMINO-2,4-DIBROMOANTHRAQUINONE 000081-49-2 | x | | | | | AMINO-2-METHYLANTHRAQUINONE, 1- 000082-28-0 | x | | | | | AMINO-3,4-DIMETHYL-3h-IMIDAZO(4,5f)QUINOLINE,2- 077094-11-2 | x | | | | | AMINO-3,8-DIMETHYL-3H-IMIDAZO(4,5-f)QUINOXALINE, -

EICG-Hot Spots: EICG Appendix C
INFORMAL REVIEW DRAFT [Note: The pre-existing regulation text is set forth below in normal type. The original proposed amendments are shown in underline to indicate additions and strikeout to indicate deletions. Additional proposed modifications are shown in double-underline to indicate additions and double-strikeout to indicate deletions. The square brackets “[ ]” are used to indicate minor adjustments to text (e.g., page numbers and adoption dates) that will be updated upon adoption of the proposed amendments.] APPENDIX C FACILITY GUIDELINE INDEX (FACILITY "LOOK-UP" TABLE) INFORMAL REVIEW DRAFT This Page Intentionally Left Blank INFORMAL REVIEW DRAFT APPENDIX C - I RESPONSIBILITIES OF ALL FACILITIES INFORMAL REVIEW DRAFT NOTES FOR APPENDIX CFACILITY GUIDELINE INDEX APPENDIX C-I RESPONSIBILITIES OF ALL FACILITIES NOTHING IN THIS APPENDIX SHALL BE CONSTRUED AS REQUIRING THAT SOURCE TESTING BE CONDUCTED FOR SUBSTANCES SET FORTH IN THIS APPENDIX. FURTHER, IN CASES WHERE A SUBSTANCE SET FORTH HEREIN IS NOT PRESENT AT A PARTICULAR FACILITY, THE FACILITY OPERATOR SHALL NOT ATTEMPT TO QUANTIFY THE EMISSIONS OF SUCH SUBSTANCE, BUT SHALL PROVIDE ADEQUATE DOCUMENTATION TO DEMONSTRATE TO THE DISTRICT THAT THE POSSIBLE PRESENCE OF THE SUBSTANCE AT THE FACILITY HAS BEEN ADDRESSED AND THAT THERE ARE NO EMISSIONS OF THE SUBSTANCE FOR SPECIFIED REASONS. Substances emitted by a particular device or process may not be limited to those listed in this Facility Guideline Index. THIS APPENDIX IS NOT AN EXHAUSTIVE LIST. ALL FACILITIES ARE RESPONSIBLE FOR IDENTIFYING AND ACCOUNTING FOR ANY LISTED SUBSTANCE USED, MANUFACTURED, FORMULATED, OR RELEASED. This Facility Guideline Index is arranged in alphabetical order. The first part of the index, Appendix C-I, lists devices common to many industries and the second part of the index, Appendix C-II, lists industry types. -

Chemical Names and CAS Numbers Final
Chemical Abstract Chemical Formula Chemical Name Service (CAS) Number C3H8O 1‐propanol C4H7BrO2 2‐bromobutyric acid 80‐58‐0 GeH3COOH 2‐germaacetic acid C4H10 2‐methylpropane 75‐28‐5 C3H8O 2‐propanol 67‐63‐0 C6H10O3 4‐acetylbutyric acid 448671 C4H7BrO2 4‐bromobutyric acid 2623‐87‐2 CH3CHO acetaldehyde CH3CONH2 acetamide C8H9NO2 acetaminophen 103‐90‐2 − C2H3O2 acetate ion − CH3COO acetate ion C2H4O2 acetic acid 64‐19‐7 CH3COOH acetic acid (CH3)2CO acetone CH3COCl acetyl chloride C2H2 acetylene 74‐86‐2 HCCH acetylene C9H8O4 acetylsalicylic acid 50‐78‐2 H2C(CH)CN acrylonitrile C3H7NO2 Ala C3H7NO2 alanine 56‐41‐7 NaAlSi3O3 albite AlSb aluminium antimonide 25152‐52‐7 AlAs aluminium arsenide 22831‐42‐1 AlBO2 aluminium borate 61279‐70‐7 AlBO aluminium boron oxide 12041‐48‐4 AlBr3 aluminium bromide 7727‐15‐3 AlBr3•6H2O aluminium bromide hexahydrate 2149397 AlCl4Cs aluminium caesium tetrachloride 17992‐03‐9 AlCl3 aluminium chloride (anhydrous) 7446‐70‐0 AlCl3•6H2O aluminium chloride hexahydrate 7784‐13‐6 AlClO aluminium chloride oxide 13596‐11‐7 AlB2 aluminium diboride 12041‐50‐8 AlF2 aluminium difluoride 13569‐23‐8 AlF2O aluminium difluoride oxide 38344‐66‐0 AlB12 aluminium dodecaboride 12041‐54‐2 Al2F6 aluminium fluoride 17949‐86‐9 AlF3 aluminium fluoride 7784‐18‐1 Al(CHO2)3 aluminium formate 7360‐53‐4 1 of 75 Chemical Abstract Chemical Formula Chemical Name Service (CAS) Number Al(OH)3 aluminium hydroxide 21645‐51‐2 Al2I6 aluminium iodide 18898‐35‐6 AlI3 aluminium iodide 7784‐23‐8 AlBr aluminium monobromide 22359‐97‐3 AlCl aluminium monochloride