Multiple Host Shifts Between Distantly Related Plants, Juglandaceae And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Synteny Analysis in Rosids with a Walnut Physical Map Reveals Slow Genome Evolution in Long-Lived Woody Perennials Ming-Cheng Luo1, Frank M
Luo et al. BMC Genomics (2015) 16:707 DOI 10.1186/s12864-015-1906-5 RESEARCH ARTICLE Open Access Synteny analysis in Rosids with a walnut physical map reveals slow genome evolution in long-lived woody perennials Ming-Cheng Luo1, Frank M. You2, Pingchuan Li2, Ji-Rui Wang1,4, Tingting Zhu1, Abhaya M. Dandekar1, Charles A. Leslie1, Mallikarjuna Aradhya3, Patrick E. McGuire1 and Jan Dvorak1* Abstract Background: Mutations often accompany DNA replication. Since there may be fewer cell cycles per year in the germlines of long-lived than short-lived angiosperms, the genomes of long-lived angiosperms may be diverging more slowly than those of short-lived angiosperms. Here we test this hypothesis. Results: We first constructed a genetic map for walnut, a woody perennial. All linkage groups were short, and recombination rates were greatly reduced in the centromeric regions. We then used the genetic map to construct a walnut bacterial artificial chromosome (BAC) clone-based physical map, which contained 15,203 exonic BAC-end sequences, and quantified with it synteny between the walnut genome and genomes of three long-lived woody perennials, Vitis vinifera, Populus trichocarpa,andMalus domestica, and three short-lived herbs, Cucumis sativus, Medicago truncatula, and Fragaria vesca. Each measure of synteny we used showed that the genomes of woody perennials were less diverged from the walnut genome than those of herbs. We also estimated the nucleotide substitution rate at silent codon positions in the walnut lineage. It was one-fifth and one-sixth of published nucleotide substitution rates in the Medicago and Arabidopsis lineages, respectively. We uncovered a whole-genome duplication in the walnut lineage, dated it to the neighborhood of the Cretaceous-Tertiary boundary, and allocated the 16 walnut chromosomes into eight homoeologous pairs. -

Characterization of the Complete Chloroplast Genome of Platycarya Strobilacea (Juglandaceae)
Conservation Genet Resour (2017) 9:79–81 DOI 10.1007/s12686-016-0624-x TECHNICAL NOTE Characterization of the complete chloroplast genome of Platycarya strobilacea (Juglandaceae) Jing Yan1 · Kai Han1 · Shuyun Zeng1 · Peng Zhao1 · Keith Woeste2 · Jianfang Li1 · Zhan-Lin Liu1 Received: 1 September 2016 / Accepted: 4 October 2016 / Published online: 6 October 2016 © Springer Science+Business Media Dordrecht 2016 Abstract. The whole chloroplast genome (cp genome) The overall AT content of the cp genome is 64 %, and the sequence of Platycarya strobilacea was characterized corresponding values of the LSC, SSC and IR regions are from Illumina pair-end sequencing data. The complete cp 66.4, 70.1 and 57.5 %, respectively. Phylogenetic analysis genome was 160,994 bp in length and contained a large confirmed the placement of P. strobilacea near to Juglans. single copy region (LSC) of 90,225 bp and a small single copy region (SSC) of 18,371 bp, which were separated Keywords Platycarya strobilacea · Hickory · China · by a pair of inverted repeat regions (IRs, 26,199 bp). The Conservation · Chloroplast genome genome contained 130 genes, including 85 protein-coding genes (80 PCG species), 36 tRNA genes (29 tRNA spe- cies) and 8 ribosomal RNA genes (4 rRNA species). Most Platycarya strobilacea, the only species in the monotypic genes occur as a single copy, but 15 genes are duplicated. genus Platycarya (Juglandaceae), mainly grows as scat- tered individuals in evergreen forests of South China (Chen et al. 2012). As one of main components of the local vegeta- tion, this species plays a key role in the forest ecosystems. -

Wingnut (Juglandaceae)
83 Wingnut (Juglandaceae) as a new generic host for Pityophthorus juglandis (Coleoptera: Curculionidae) and the thousand cankers disease pathogen, Geosmithia morbida (Ascomycota: Hypocreales) Stacy M. Hishinuma, Paul L. Dallara, Mohammad A. Yaghmour, Marcelo M. Zerillo, Corwin M. Parker, Tatiana V. Roubtsova, Tivonne L. Nguyen, Ned A. Tisserat, Richard M. Bostock, Mary L. Flint, Steven J. Seybold1 Abstract—The walnut twig beetle (WTB), Pityophthorus juglandis Blackman (Coleoptera: Curculionidae), vectors a fungus, Geosmithia morbida Kolařík, Freeland, Utley, and Tisserat (Ascomycota: Hypocreales), which colonises and kills the phloem of walnut and butternut trees, Juglans Linnaeus (Juglandaceae). Over the past two decades, this condition, known as thousand cankers disease (TCD), has led to the widespread mortality of Juglans species in the United States of America. Recently the beetle and pathogen were discovered on several Juglans species in northern Italy. Little is known about the extra-generic extent of host acceptability and suitability for the WTB. We report the occurrence of both the WTB and G. morbida in three species of wingnut, Pterocarya fraxinifolia Spach, Pterocarya rhoifolia Siebold and Zuccarini, and Pterocarya stenoptera de Candolle (Juglandaceae) growing in the United States Department of Agriculture-Agricultural Research Service, National Clonal Germplasm Repository collection in northern California (NCGR) and in the Los Angeles County Arboretum and Botanic Garden in southern California, United States of America. In two instances (once in P. stenoptera and once in P. fraxinifolia) teneral (i.e., brood) adult WTB emerged and were collected more than four months after infested branch sections had been collected in the field. Koch’s postulates were satisfied with an isolate of G. -

(Juglandaceae) in the Early Miocene of Panama
CHAPTER 7 FRUITS OF OREOMUNNEA (JUGLANDACEAE) IN THE EARLY MIOCENE OF PANAMA Fabiany Herrera, Steven R. Manchester,* Rebecca Koll, and Carlos Jaramillo ABSTRACT Permineralized fruits of Juglandaceae have been recovered from the Early Miocene Cucaracha Formation exposed in excavations of the Panama Canal. Transverse sections and peels reveal that the nuts are subdivided into eight chambers at the base, and the nutshell and septa are composed mainly of fibers. Together, these features are found only in the Neotropical juglandaceous clade composed of extant Alfaroa Standl. and Oreomunnea Oerst. Based on the small nut size and rela- tively thin wall and septa, these nuts are assigned to Oreomunnea, which is distinguished from extant Alfaroa by its winged fruits adapted for wind dispersal. A. Graham first reported pollen grains of the Alfaroa/Oreomunnea type from the Miocene of Panama. The new fossil fruits of the Cucaracha Formation are the earliest macrofossil record of Oreomunnea, enhancing our under- standing of the phytogeographic history of Juglandaceae in Central America. If this newly de- scribed fossil species, O. grahamii Manch. & Herrera, grew in similar ecological settings to those of its modern counterparts, the fossils may be an indicator of the medium-high paleotopographic relief of Panama during the Early Miocene. Key words: Alfaroa, fossils, fruits, Juglandaceae, Miocene, Oreomunnea. The Juglandaceae have an excellent fossil rec- Neotropical genera Alfaroa Standl. and Oreo- ord in the Northern Hemisphere, well represented munnea Oerst. have mostly eluded detection in by pollen, leaves, wood, catkins, and fruits. This the macrofossil record. record includes most of the extant genera, plus a Ten genera in two subfamilies are recognized few extinct ones in the Paleogene (Manchester, for the Juglandaceae (Manos & Stone, 2001; 1987, 1989; Elliot et al., 2006). -

Inflorescence Dimorphism, Heterodichogamy and Thrips
Annals of Botany 113: 467–476, 2014 doi:10.1093/aob/mct278, available online at www.aob.oxfordjournals.org Inflorescence dimorphism, heterodichogamy and thrips pollination in Platycarya strobilacea (Juglandaceae) Tatsundo Fukuhara* and Shin-ichiro Tokumaru Faculty of Education, Fukuoka University of Education, 1-1 Akama-Bunkyo-machi, Munakata, Fukuoka, Japan * For correspondence. E-mail [email protected] Received: 22 July 2013 Returned for revision: 11 September 2013 Accepted: 14 October 2013 Published electronically: 3 December 2013 † Background and Aims Unlike other taxa in Juglandaceae or in closely related families, which are anemophilous, Platycarya strobilacea has been suggested to be entomophilous. In Juglandaceae, Juglans and Carya show hetero- dichogamy, a reproductive strategy in which two morphs coexist in a population and undergo synchronous reciprocal sex changes. However, there has been no study focusing on heterodichogamy in the other six or seven genera, includ- ing Platycarya. † Methods Inflorescence architecture, sexual expression and pollination biology were examined in a P. strobilacea population in Japan. Flowering phenology was monitored daily for 24 trees in 2008 and 27 in 2009. Flower visitors and inhabitants were recorded or collected from different sexes and stages. † Key results The population of P. strobilacea showed heterodichogamous phenology with protogynous and duodi- chogamous–protandrous morphs. This dimorphism in dichogamy was associated with distinct inflorescence morph- ologies.Thrips pollination was suggested bythe frequent presence of thrips withattached pollen grains,the scarcityof other insect visitors, the synchronicity of thrips number in male spikes with the maturation of female flowers, and morphological characters shared with previously reported thrips-pollinated plants. Male spikes went through two consecutive stages: bright yellow and strong-scented M1 stage, and brownish and little-scented M2 stage. -

Fragrant Annuals Fragrant Annuals
TheThe AmericanAmerican GARDENERGARDENER® TheThe MagazineMagazine ofof thethe AAmericanmerican HorticulturalHorticultural SocietySociety JanuaryJanuary // FebruaryFebruary 20112011 New Plants for 2011 Unusual Trees with Garden Potential The AHS’s River Farm: A Center of Horticulture Fragrant Annuals Legacies assume many forms hether making estate plans, considering W year-end giving, honoring a loved one or planting a tree, the legacies of tomorrow are created today. Please remember the American Horticultural Society when making your estate and charitable giving plans. Together we can leave a legacy of a greener, healthier, more beautiful America. For more information on including the AHS in your estate planning and charitable giving, or to make a gift to honor or remember a loved one, please contact Courtney Capstack at (703) 768-5700 ext. 127. Making America a Nation of Gardeners, a Land of Gardens contents Volume 90, Number 1 . January / February 2011 FEATURES DEPARTMENTS 5 NOTES FROM RIVER FARM 6 MEMBERS’ FORUM 8 NEWS FROM THE AHS 2011 Seed Exchange catalog online for AHS members, new AHS Travel Study Program destinations, AHS forms partnership with Northeast garden symposium, registration open for 10th annual America in Bloom Contest, 2011 EPCOT International Flower & Garden Festival, Colonial Williamsburg Garden Symposium, TGOA-MGCA garden photography competition opens. 40 GARDEN SOLUTIONS Plant expert Scott Aker offers a holistic approach to solving common problems. 42 HOMEGROWN HARVEST page 28 Easy-to-grow parsley. 44 GARDENER’S NOTEBOOK Enlightened ways to NEW PLANTS FOR 2011 BY JANE BERGER 12 control powdery mildew, Edible, compact, upright, and colorful are the themes of this beating bugs with plant year’s new plant introductions. -
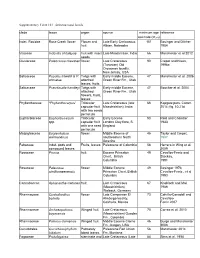
Supplementary Table 10.1. Selected Rosid Fossils Clade Taxon Organ Source Minimum Age Reference Estimate (Mya) Indet
Supplementary Table 10.1. Selected rosid fossils clade taxon organ source minimum age reference estimate (Mya) Indet. Rosidae Rose Creek flower Flower and Late Early Cretaceous 101 Basinger and Dilcher fruit Albian, Nebraska 1984 Vitaceae Indovitis chitaleyae fruit with intact Late Maastrictian, India 66 Manchester et al 2012 seeds Clusiaceae Paleoclusia chevalieri flower Late Cretaceous 90 Crepet and Nixon, (Turonian) Old 1998a Crossman locality, New Jersey, USA Salicaceae Populus tidwellii & P. Twigs with Early middle Eocene, 47 Manchester et al. 2006 wilmatae attached Green River Fm., Utah leaves, fruits Salicaceae Pseudosalix handleyi Twigs with Early middle Eocene, 47 Boucher et al. 2004 attached Green River Fm., Utah flowers, fruits, leaves Phyllanthaceae “Phyllanthocarpus” Trilocular Late Cretaceous (late 66 Kapgate pers. Comm. capsular fruit Maastrichtian), India 2014; fig. 10.21d with two seeds per locule Euphorbiaceae Euphorbiocarpon Trilocular Early Eocene 50 Reid and Chandler spp. capsular fruit London Clay flora, S. 1933 with one seed England per locule Malpighiaceae Eoglandulosa flower Middle Eocene of 45 Taylor and Crepet, warmanensis southeastern North 1987 America Fabaceae Indet. pods and Fruits, leaves Paleocene of Colombia 58 Herrera in Wing et al. compound leaves 2009 Rosaceae Prunus fruit Eocene Princeton 49 Cevallos-Ferriz and Chert, British Stockey, Columbia 1991 Rosaceae Paleorosa flower Middle Eocene 49 Basinger 1976; similkameenensis Princeton Chert, British Cevallos-Ferriz , et al Columbia 1993 Cannabaceae Aphananthe cretacea fruit Late Cretaceous 67 Knobloch and Mai (Maastrichtian) 1986 Walbeck, Germany Rhamnaceae Coahuilanthus flower Late Campanian El 73 Calvillo-Canadell and belinda Almácigo locality, Cevallos- Coahuila, Mexico Ferriz 2007 Rhamnaceae Archaeopaliurus Winged fruit Late Cretaceous 70 Correa et al. -

Whole Genome Based Insights Into the Phylogeny and Evolution of the Juglandaceae
Whole Genome based Insights into the Phylogeny and Evolution of the Juglandaceae Huijuan Zhou Northwest A&F University: Northwest Agriculture and Forestry University Yiheng Hu Northwestern University Aziz Ebrahimi Purdue University Peiliang Liu Northwestern University Keith Woeste Purdue University Shuoxin Zhang Northwest A&F University: Northwest Agriculture and Forestry University Peng Zhao ( [email protected] ) Northwest University https://orcid.org/0000-0003-3033-6982 Research article Keywords: Diversication, Divergence time, Genome, Juglandaceae, Phylogenomics, Plastome Posted Date: May 24th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-495294/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/23 Abstract Background: The walnut family (Juglandaceae) contains commercially important woody trees commonly called walnut, wingnut, pecan and hickory. Phylogenetic relationships in the Juglandaceae are problematic, and their historical diversication has not been claried, in part because of low phylogenetic resolution and/or insucient marker variability. Results: We reconstructed the backbone phylogenetic relationships of Juglandaceae using organelle and nuclear genome data from 27 species. The divergence time of Juglandaceae was estimated to be 78.7 Mya. The major lineages diversied in warm and dry habitats during the mid-Paleocene and early Eocene. The plastid, mitochondrial, and nuclear phylogenetic analyses all revealed three subfamilies, i.e., Juglandoideae, Engelhardioideae, Rhoipteleoideae. Five genera of Juglandoideae were strongly supported. Juglandaceae were estimated to have originated during the late Cretaceous, while Juglandoideae were estimated to have originated during the Paleocene, with evidence for rapid diversication events during several glacial and geological periods. The phylogenetic analyses of organelle sequences and nuclear genome yielded highly supported incongruence positions for J. -

Supplementary Material
Xiang et al., Page S1 Supporting Information Fig. S1. Examples of the diversity of diaspore shapes in Fagales. Fig. S2. Cladogram of Fagales obtained from the 5-marker data set. Fig. S3. Chronogram of Fagales obtained from analysis of the 5-marker data set in BEAST. Fig. S4. Time scale of major fagalean divergence events during the past 105 Ma. Fig. S5. Confidence intervals of expected clade diversity (log scale) according to age of stem group. Fig. S6. Evolution of diaspores types in Fagales with BiSSE model. Fig. S7. Evolution of diaspores types in Fagales with Mk1 model. Fig. S8. Evolution of dispersal modes in Fagales with MuSSE model. Fig. S9. Evolution of dispersal modes in Fagales with Mk1 model. Fig. S10. Reconstruction of pollination syndromes in Fagales with BiSSE model. Fig. S11. Reconstruction of pollination syndromes in Fagales with Mk1 model. Fig. S12. Reconstruction of habitat shifts in Fagales with MuSSE model. Fig. S13. Reconstruction of habitat shifts in Fagales with Mk1 model. Fig. S14. Stratigraphy of fossil fagalean genera. Table S1 Genera of Fagales indicating the number of recognized and sampled species, nut sizes, habits, pollination modes, and geographic distributions. Table S2 List of taxa included in this study, sources of plant material, and GenBank accession numbers. Table S3 Primers used for amplification and sequencing in this study. Table S4 Fossil age constraints utilized in this study of Fagales diversification. Table S5 Fossil fruits reviewed in this study. Xiang et al., Page S2 Table S6 Statistics from the analyses of the various data sets. Table S7 Estimated ages for all families and genera of Fagales using BEAST. -

STOR ® Scott L
The Platycarya Perplex and the Evolution of the Juglandaceae STOR ® Scott L. Wing; Leo J. Hickey American Journal of Botany, Vol. 71, No. 3. (Mar., 1984), pp. 388-411. Stable URL: http://links.jstor.org/sici?sici=0002-9122%28198403%2971%3A3%3C388%3ATPPATE%3E2.0.CO%3B2-A American Journal of Botany is currently published by Botanical Society of America. Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/journals/botsam.html. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is an independent not-for-profit organization dedicated to and preserving a digital archive of scholarly journals. For more information regarding JSTOR, please contact [email protected]. http ://www.j stor.org Fri Dec 15 12:53:47 2006 Amer. J. Bot. 71(3): 388-411. 1984. THE PLATYCARYA PERPLEX AND THE EVOLUTION OF THE JUGLANDACEAE1 SCOTT L. WING AND LEO J. HICKEY U.S. Geological Survey, E-501 National Museum of Natural History, Washington, DC. -

BMC Evolutionary Biology Biomed Central
BMC Evolutionary Biology BioMed Central Research article Open Access Mitochondrial matR sequences help to resolve deep phylogenetic relationships in rosids Xin-Yu Zhu1,2, Mark W Chase3, Yin-Long Qiu4, Hong-Zhi Kong1, David L Dilcher5, Jian-Hua Li6 and Zhi-Duan Chen*1 Address: 1State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, the Chinese Academy of Sciences, Beijing 100093, China, 2Graduate University of the Chinese Academy of Sciences, Beijing 100039, China, 3Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK, 4Department of Ecology & Evolutionary Biology, The University Herbarium, University of Michigan, Ann Arbor, MI 48108-1048, USA, 5Florida Museum of Natural History, University of Florida, Gainesville, FL 32611-7800, USA and 6Arnold Arboretum of Harvard University, 125 Arborway, Jamaica Plain, MA 02130, USA Email: Xin-Yu Zhu - [email protected]; Mark W Chase - [email protected]; Yin-Long Qiu - [email protected]; Hong- Zhi Kong - [email protected]; David L Dilcher - [email protected]; Jian-Hua Li - [email protected]; Zhi- Duan Chen* - [email protected] * Corresponding author Published: 10 November 2007 Received: 19 June 2007 Accepted: 10 November 2007 BMC Evolutionary Biology 2007, 7:217 doi:10.1186/1471-2148-7-217 This article is available from: http://www.biomedcentral.com/1471-2148/7/217 © 2007 Zhu et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. -

National Park Service Paleontological Research
169 NPS Fossil National Park Service Resources Paleontological Research Edited by Vincent L. Santucci and Lindsay McClelland Technical Report NPS/NRGRD/GRDTR-98/01 United States Department of the Interior•National Park Service•Geological Resource Division 167 To the Volunteers and Interns of the National Park Service iii 168 TECHNICAL REPORT NPS/NRGRD/GRDTR-98/1 Copies of this report are available from the editors. Geological Resources Division 12795 West Alameda Parkway Academy Place, Room 480 Lakewood, CO 80227 Please refer to: National Park Service D-1308 (October 1998). Cover Illustration Life-reconstruction of Triassic bee nests in a conifer, Araucarioxylon arizonicum. NATIONAL PARK SERVICE PALEONTOLOGICAL RESEARCH EDITED BY VINCENT L. SANTUCCI FOSSIL BUTTE NATIONAL MONUMNET P.O. BOX 592 KEMMERER, WY 83101 AND LINDSAY MCCLELLAND NATIONAL PARK SERVICE ROOM 3229–MAIN INTERIOR 1849 C STREET, N.W. WASHINGTON, D.C. 20240–0001 Technical Report NPS/NRGRD/GRDTR-98/01 October 1998 FORMATTING AND TECHNICAL REVIEW BY ARVID AASE FOSSIL BUTTE NATIONAL MONUMENT P. O . B OX 592 KEMMERER, WY 83101 164 165 CONTENTS INTRODUCTION ...............................................................................................................................................................................iii AGATE FOSSIL BEDS NATIONAL MONUMENT Additions and Comments on the Fossil Birds of Agate Fossil Beds National Monument, Sioux County, Nebraska Robert M. Chandler ..........................................................................................................................................................................