<I>Euphorbiaceae</I>
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cara Membaca Informasi Daftar Jenis Tumbuhan
Dilarang mereproduksi atau memperbanyak seluruh atau sebagian dari buku ini dalam bentuk atau cara apa pun tanpa izin tertulis dari penerbit. © Hak cipta dilindungi oleh Undang-Undang No. 28 Tahun 2014 All Rights Reserved Rugayah Siti Sunarti Diah Sulistiarini Arief Hidayat Mulyati Rahayu LIPI Press © 2015 Lembaga Ilmu Pengetahuan Indonesia (LIPI) Pusat Penelitian Biologi Katalog dalam Terbitan (KDT) Daftar Jenis Tumbuhan di Pulau Wawonii, Sulawesi Tenggara/ Rugayah, Siti Sunarti, Diah Sulistiarini, Arief Hidayat, dan Mulyati Rahayu– Jakarta: LIPI Press, 2015. xvii + 363; 14,8 x 21 cm ISBN 978-979-799-845-5 1. Daftar Jenis 2. Tumbuhan 3. Pulau Wawonii 158 Copy editor : Kamariah Tambunan Proofreader : Fadly S. dan Risma Wahyu H. Penata isi : Astuti K. dan Ariadni Desainer Sampul : Dhevi E.I.R. Mahelingga Cetakan Pertama : Desember 2015 Diterbitkan oleh: LIPI Press, anggota Ikapi Jln. Gondangdia Lama 39, Menteng, Jakarta 10350 Telp. (021) 314 0228, 314 6942. Faks. (021) 314 4591 E-mail: [email protected] Website: penerbit.lipi.go.id LIPI Press @lipi_press DAFTAR ISI DAFTAR GAMBAR ............................................................................. vii PENGANTAR PENERBIT .................................................................. xi KATA PENGANTAR ............................................................................ xiii PRAKATA ............................................................................................. xv PENDAHULUAN ............................................................................... -

Arborescent Angiosperms of Mundanthurai Range in The
Check List 8(5): 951–962, 2012 © 2012 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution Arborescent Angiosperms of Mundanthurai Range in PECIES S the Kalakad-Mundanthurai Tiger Reserve (KMTR) of the OF southern Western Ghats, India ISTS L Paulraj Selva Singh Richard 1* and Selvaraj Abraham Muthukumar 2 1 Madras Christian College, Department of Botany, Chennai – 600 059, Tamil Nadu, India. 2 St. John’s College, Department of Botany, Tirunelveli, 627 002, Tamil Nadu, India. [email protected] * Corresponding author. E-mail: Abstract: The present study was carried out to document the diversity of arborescent angiosperm taxa of Mundanthurai representingRange in the 175Kalakad-Mundanthurai genera in 65 families Tiger were Reserve recorded. (KMTR) The most of the speciose southern families Western are Euphorbiaceae Ghats in India. (27 During spp.), the Rubiaceae floristic survey carried out from January 2008 to December 2010, a total of 247 species and intraspecific taxa of trees and shrubs to this region which includes Agasthiyamalaia pauciflora, Elaeocarpus venustus, Garcinia travancorica, Gluta travancorica, (17Goniothalamus spp.), Myrtaceae rhynchantherus, (14 spp.), Lauraceae Homalium (13 travancoricum, spp.) and Annonaceae Homaium (11 jainii, spp.). OropheaOf the 247 uniflora, taxa, 27 Phlogacanthus species are endemic albiflorus, only Polyalthia shendurunii, Symplocos macrocarpa and Symplocos sessilis . This clearly signifies that this range is relevant to the conservation of the local flora. Introduction India for conserving global biological diversity and also The Western Ghats is one of the biodiversity hotspots declared as Regional Centre of Endemism in the Indian of the world (Myers et al. -

Discovering Karima (Euphorbiaceae), a New Crotonoid Genus from West Tropical Africa Long Hidden Within Croton
RESEARCH ARTICLE Discovering Karima (Euphorbiaceae), a New Crotonoid Genus from West Tropical Africa Long Hidden within Croton Martin Cheek1*, Gill Challen1, Aiah Lebbie2, Hannah Banks1, Patricia Barberá3, Ricarda Riina3* 1 Science Department, Royal Botanic Gardens, Kew, Surrey, United Kingdom, 2 National Herbarium of Sierra Leone, Dept. of Biological Sciences, Njala University, PMB, Freetown, Sierra Leone, 3 Department of Biodiversity and Conservation, Real Jardín Botánico, RJB-CSIC, Plaza de Murillo, Madrid, Spain * [email protected] (MC); [email protected] (RR) Abstract Croton scarciesii (Euphorbiaceae-Crotonoideae), a rheophytic shrub from West Africa, is OPEN ACCESS shown to have been misplaced in Croton for 120 years, having none of the diagnostic char- Citation: Cheek M, Challen G, Lebbie A, Banks H, acters of that genus, but rather a set of characters present in no known genus of the family. Barberá P, Riina R (2016) Discovering Karima Pollen analysis shows that the new genus Karima belongs to the inaperturate crotonoid (Euphorbiaceae), a New Crotonoid Genus from West Tropical Africa Long Hidden within Croton. PLoS group. Analysis of a concatenated molecular dataset combining trnL-F and rbcL sequences ONE 11(4): e0152110. doi:10.1371/journal. positioned Karima as sister to Neoholstia from south eastern tropical Africa in a well-sup- pone.0152110 ported clade comprised of genera of subtribes Grosserineae and Neoboutonieae of the ina- Editor: Nico Cellinese, University of Florida, UNITED perturate crotonoid genera. Several morphological characters support the relationship of STATES Karima with Neoholstia, yet separation is merited by numerous characters usually associ- Received: January 5, 2016 ated with generic rank in Euphorbiaceae. -

Labiatae) in India (Ph.D
Scientific Journey (from 1988 to till date) V. SAMPATH KUMAR Scientist, CNH, BSI, Howrah Taxonomic studies on the tribe Mentheae (Labiatae) in India (Ph.D. – Calcutta University) Research scholar of CNH, BSI under ‘Flora of India’ project from 1988 to 1993. Scientific Asst. from 1993 to 1997. Botanist from 1997 to 2001 (ANRC, PortBlair). Scientist (CDRI, Lucknow) from 2001 to 2002. Scientist in BSI since Sept. 2002. Served in Hqrs., Kolkata; DRC, Hyderabad; SRC, Coimbatore; RBG, Kew (as Indian Botanical Liaison officer); presently working at CNH, Howrah. Botanist Andaman & Nicobar Regional Centre, PortBlair (1997-2001) Study of floristic & ecological aspects of Ritchie’s Archipelago (incl. Rani Jhansi Marine National Park, 1998- 2000) and Kalpong Hydro-electric project site (2000-01). The Kalpong HEP study jointly made with Dr. P.V. Sreekumar, the then Scientist of ANRC. Flora of Andaman & Nicobar Islands (vol. II) Myrtaceae to Alangiaceae (16+3 = 19 families) (co-authored by L.N. Ray and P.S.N. Rao) Alangiaceae, Apiaceae, Araliaceae, Barringtoniaceae, Begoniaceae, Cactaceae, Caricaceae, Cornaceae, Cucurbitaceae, Datiscaceae, Lecythidaceae, Lythraceae, Melastomataceae, Memecylaceae, Myrtaceae, Onagraceae, Passifloraceae, Punicaceae, and Sonneratiaceae Mangroves of Andaman & Nicobar Islands 32 species recognized out of 35 reported (all species found in Andaman group; only15 from Nicobar group). Added – Xylocarpus gangeticus (Prain) Parkinson (not included by Dagar et al. (1991) and Dagar & Singh (1999); Banerjee (2002) in his Indian account) Excluded – Bruguiera sexangula (Lour.) Poiret, Cynometra ramiflora L. (not found in India), Rhizophora x lamarckii Mont., and Sonneratia apetala Buch.-Ham. Vernacular names of A & N Island Plants The plants’ vernacular names used by the tribes of these islands along with their uses were compiled and submitted (500 pp.). -

Revision of Annesijoa, Elateriospermum and the Introduced Species of Hevea in Malesia (Euphorbiaceae)
BLUMEA 49: 425– 440 Published on 10 December 2004 doi: 10.3767/000651904X484351 REVISION OF ANNESIJOA, ELATERIOSPERMUM AND THE INTRODUCED SPECIES OF HEVEA IN MALESIA (EUPHORBIACEAE) HOANG VAN SAM1 & PETER C. vaN WELZEN2 SUMMARY Annesijoa is an endemic monotypic genus from New Guinea with as single species A. novoguineensis. Elateriospermum is also monotypic (E. tapos) and found in West Malesia. The South American genus Hevea comprises about 10 species. One species (H. brasiliensis) is presently cultivated worldwide in plantations for its rubber and has become one of the major economic products of SE Asia. Two other species, H. guianensis and H. pauciflora are sometimes present in Malesian botanical gardens. Key words: Euphorbiaceae, Annesijoa, Elateriospermum, Hevea, Malesia. INTRODUCTION Three genera are revised for Flora Malesiana, Annesijoa Pax & K. Hoffm., Elaterio spermum Blume, and Hevea Aubl. These genera are not very closely related, but they are all part of the subfamily Crotonoideae (Webster, 1994; Radcliffe-Smith, 2001), though classified in different tribes (Jatropheae, Elateriospermeae, and Micrandreae subtribe Heveinae, respectively). Typical for the Micrandreae are colporate pollen with a reticulate sexine, articulate laticifers, absent petals, and plenty of endosperm (oily in the Heveinae). The Jatropheae and Elateriospermeae share inaperculate pollen with a typical ‘crotonoid’ sexine, inarticulate laticifers, petals absent or not, and seeds with or without endosperm. They differ in several characters, the Elateriospermeae lack endosperm and have no petals, while these are present in the Jatropheae. The three genera can easily be distinguished from each other. Hevea and Elaterio spermum have white latex, whereas Annesijoa has variable latex ranging from clear to white to red. -

Key to the Families and Genera of Malesian <I>Euphorbiaceae</I> in the Wide Sense
Blumea 65, 2020: 53–60 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE https://doi.org/10.3767/blumea.2020.65.01.05 Key to the families and genera of Malesian Euphorbiaceae in the wide sense P.C. van Welzen1,2 Key words Abstract Identification keys are provided to the different families in which the Euphorbiaceae are split after APG IV. Presently, Euphorbiaceae in the strict sense, Pandaceae, Peraceae, Phyllanthaceae, Picrodendraceae and Euphorbiaceae Putranjivaceae are distinguished as distinct families. Within the families, keys to the different genera occurring in the keys Malesian area, native and introduced, are presented. The keys are to be tested and responses are very welcome. Pandaceae Peraceae Published on 3 April 2020 Phyllanthaceae Picrodendraceae Putranjivaceae INTRODUCTION KEY TO THE EUPHORBIACEOUS FAMILIES The Euphorbiaceae in the wide sense (sensu lato, s.lat.) were 1. Ovary with a single ovule per locule . 2 always a heterogeneous group without any distinct combination 1. Ovary with two ovules per locule ................... 4 of characters. The most typical features are the presence of uni- 2. Fruits drupes. Flowers of both sexes with petals . sexual simple flowers and fruits that fall apart in various carpel ...................................2. Pandaceae fragments and seeds, leaving the characteristic columella on the 2. Fruits capsules, sometimes drupes or berries, then flowers plant. However, some groups, like the Pandaceae and Putranji of both sexes lacking petals.......................3 vaceae, were morphologically already known to be quite unlike 3. Herbs, shrubs, lianas, trees, mono- or dioecious. Flowers in the rest of the Euphorbiaceae s.lat. (e.g., Radcliffe-Smith 1987). cauliflorous, ramiflorous, axillary, or terminal inflorescences The Putranjivaceae formerly formed the tribe Drypeteae in the ................................1. -

EBENACEAE Diospyros Malabarica (Descr.) Kostel. Synonyms
Mangrove Guidebook for Southeast Asia Part 2: DESCRIPTIONS – Trees & shrubs EBENACEAE 171 Diospyros malabarica (Descr.) Kostel. Synonyms : Diospyros embryopteris Pers., Diospyros embryopteris var. siamensis (Hochr.) Phengklai, Diospyros glutinosa Koenig. & Roxb., Diospyros malabarica var. malabarica, Diospyros melanoxylon Hassk., Diospyros peregrina Guerke., Diospyros peregrina f. javanica (Gaert.) Guerke., Diospyros siamensis Hochr., Diospyros siamensis Ridl., Garcinia malabarica Desr. Vernacular name(s) : River ebony, Indian persimmon, Mountain ebony, Malabar ebony (E), Komoi, Kumun (Mal.), Culiket, Klega, Kleca, Toyokuku, Makusi (Ind.), Tako suan (Thai) Description : Evergreen dioecious tree with male and female flowers occurring on different trees, medium to large sized, up to 37 m tall. The trunk may have a girth of up to 2 m and is often fluted at the base; bark is almost black. Leaves are simple, alternate, leathery, 3-6(-10) cm by 10-20(-24) cm, oblong, 6-8 side veins, sunken mid-vein, and often with a pointed tip. Leaf stalk 10-12 mm long. Young leaves are reddish. Flowers are whitish. Male flower clusters have 3-5 flowers, 4-merous, each up to 6 mm wide, with valve-like calyx lobes and 24-47 stamens. Female flowers are solitary in lead axils, 18-25 mm wide, with a 1 cm long stalk; 4- or 5-merous. Fruit is round or egg-shaped, 4-5 cm long, 3.5 cm diameter, yellowish to rusty brown, 6-8 partitions, with 3-8 seeds. Fruit is at first densely hairy, later becoming smooth; 4 calyx lobes are hairy and remain attached to fruit. Fruit pulp is glutinous. Ecology : Often cultivated in homestead gardens. -

A Taxonomic Revision of Trigonostemon (Euphorbiaceae) in Malesia
Blumea 62, 2018: 179–229 ISSN (Online) 2212-1676 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE https://doi.org/10.3767/blumea.2018.62.03.04 A taxonomic revision of Trigonostemon (Euphorbiaceae) in Malesia R.-Y. Yu1, P.C. van Welzen1,2 Key words Abstract Trigonostemon is taxonomically revised for Malesia based on herbarium collections and field observa- tions. Thirty-eight species are recognized in Malesia, of which four of uncertain status and four newly described. Euphorbiaceae The previous infrageneric classifications are briefly reviewed, but none is accepted. Some useful characters are Malesia discussed. An identification key, nomenclature, descriptions, typification, geographic distributions and taxonomic morphological revision notes are provided. taxonomy Trigonostemon Published on 30 January 2018 INTRODUCTION of stamens, only the inner 2 whorls connate). Later, two other genera, Dimorphocalyx Thwaites (1861; staminate flowers Trigonostemon Blume is a genus in the Euphorbiaceae sub- with 2 whorls of stamens, only the inner whorl connate) and family Crotonoideae (phylogenetically supported based on Tylosepalum Kurz ex Teijsm. & Binn. (Teijsmann & Binnen dijk molecular data by Wurdack et al. 2005). Within the Croto- 1864; 3 connate stamens and a gland on the sepals), were noideae it is traditionally classified in tribe Trigonostemoneae described. All of them were morphologically rather similar (Webster 1975, 1994, Radcliffe-Smith 2001) or tribe Codiaeae and this triggered a discussion about the circumscription and subtribe Trigonostemoniae (Webster 2014), but none of these infrageneric classification of Trigonostemon. treatments has been confirmed by a molecular phylogeny. The Müller Argoviensis (1865, 1866) considered Trigonostemon in genus contains about 60 species ranging from India to China, a wide sense, a genus that did not only include species with throughout mainland SE Asia and Malesia to NE Australia and one whorl of 3 or 5 connate stamens, as Blume (1825) defined the W Pacific (Govaerts et al. -

Vegetation Recovery of Logged-Over Dipterocarp Forests in Central Kalimantan, Indonesia
Floresta e Ambiente 2019; 26(3): e20171239 https://doi.org/10.1590/2179-8087.123917 ISSN 2179-8087 (online) Original Article Conservation of Nature Vegetation Recovery of Logged-over Dipterocarp Forests In Central Kalimantan, Indonesia Prijanto Pamoengkas1 , Ayi Zamzam1 , Aji Dwisutono1 1Department of Silviculture, Faculty of Forestry, Bogor Agricultural University – IPB, Bogor/West Java, Indonesia ABSTRACT Forest utilization usually has an impact on changes in forest structure and species composition. The species of trees selected and ecosystem management system that refer to biodiversity characteristics will be explained by better knowledge of Functional Species Group (FSG). The purpose of this study was to determine the composition of vegetation recovery based on FSG in production forest managed with silvicultural system known as Selective Cutting and Line Planting System (SCLP) and undisturbed forest known as Germplasm Preservation Areas (GPCA) in Central Kalimantan, Indonesia. The results showed that the proportion of climax group in the entire observation plots was greater than pioneer species group, i.e. climax 100 species and pioneer 59 species. Stand structure both climax and pioneer species group has inverse J shape curve. It means that the species composition of logged-over forest is still in balance condition, characterized with high index of diversity (H’ > 3). Keywords: functional species group, selective cutting, species composition, stand structure, biodiversity. Creative Commons License. All the contents of this journal, except where otherwise noted, is licensed under a Creative Commons Attribution License. 2/11 Pamoengkas P, Zamzam A, Dwisutono A Floresta e Ambiente 2019; 26(3): e20171239 1. INTRODUCTION of utilizing dipterocarps species using SCLP is for managing natural forests to increase the productivity Undisturbed lowland dipterocarp forests have clean of tropical rainforest and maintain the biodiversity. -
A Revision of the Malesian Genus Blumeodendron (Euphorbiaceae)
Blumea 61, 2016: 64–82 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE http://dx.doi.org/10.3767/000651916X691547 A revision of the Malesian genus Blumeodendron (Euphorbiaceae) M.A.D. Ottens-Treurniet1, P.C. van Welzen1,2 Key words Abstract The genus Blumeodendron (Euphorbiaceae) comprises nine species and is distributed from the Andaman Islands and Myanmar in the west through Malesia to the Bismarck Archipelago in the east. Two species are newly Blumeodendron described, B. gesinus, with ridged fruits, and B. endocarpum, with a thickened endocarp around each seed. Three Euphorbiaceae former synonyms, B. novoguineense (new name for Bennettia papuanum), B. philippinense and B. subrotundifolium, Malesia are reinstated as species. Two important characters were newly discovered, the presence of small lepidote hairs revision in all species and the indehiscent fruit of B. endocarpum. taxonomy Published on 29 April 2016 INTRODUCTION The variation within Blumeodendron is very high, especially in the former circumscription of the two widespread species. It is Blumeodendron (Müll.Arg.) Kurz is a tree genus in the Euphor typically a genus where it takes time to become familiar with biaceae, which ranges from the Andaman Islands and Myanmar the species before a sensible delimitation can be made. As a in the west through Malesia to the Bismarck Archipelago in the result, the species circumscriptions have changed dramatically. east. Blumeodendron belongs to the subfamily Acalyphoideae, The definition of the widespread B. kurzii and B. tokbrai is much tribe Pycnocomeae, subtribe Blumeodendrinae (Webster 1994, more restricted as B. borneense Pax & K.Hoffm. (with B. con 2014, Radcliffe-Smith 2001) and is part of clade A1 of the core color Gage as synonym), B. -

Tree Types of the World Map
Abarema abbottii-Abarema acreana-Abarema adenophora-Abarema alexandri-Abarema asplenifolia-Abarema auriculata-Abarema barbouriana-Abarema barnebyana-Abarema brachystachya-Abarema callejasii-Abarema campestris-Abarema centiflora-Abarema cochleata-Abarema cochliocarpos-Abarema commutata-Abarema curvicarpa-Abarema ferruginea-Abarema filamentosa-Abarema floribunda-Abarema gallorum-Abarema ganymedea-Abarema glauca-Abarema idiopoda-Abarema josephi-Abarema jupunba-Abarema killipii-Abarema laeta-Abarema langsdorffii-Abarema lehmannii-Abarema leucophylla-Abarema levelii-Abarema limae-Abarema longipedunculata-Abarema macradenia-Abarema maestrensis-Abarema mataybifolia-Abarema microcalyx-Abarema nipensis-Abarema obovalis-Abarema obovata-Abarema oppositifolia-Abarema oxyphyllidia-Abarema piresii-Abarema racemiflora-Abarema turbinata-Abarema villifera-Abarema villosa-Abarema zolleriana-Abatia mexicana-Abatia parviflora-Abatia rugosa-Abatia spicata-Abelia corymbosa-Abeliophyllum distichum-Abies alba-Abies amabilis-Abies balsamea-Abies beshanzuensis-Abies bracteata-Abies cephalonica-Abies chensiensis-Abies cilicica-Abies concolor-Abies delavayi-Abies densa-Abies durangensis-Abies fabri-Abies fanjingshanensis-Abies fargesii-Abies firma-Abies forrestii-Abies fraseri-Abies grandis-Abies guatemalensis-Abies hickelii-Abies hidalgensis-Abies holophylla-Abies homolepis-Abies jaliscana-Abies kawakamii-Abies koreana-Abies lasiocarpa-Abies magnifica-Abies mariesii-Abies nebrodensis-Abies nephrolepis-Abies nordmanniana-Abies numidica-Abies pindrow-Abies pinsapo-Abies -
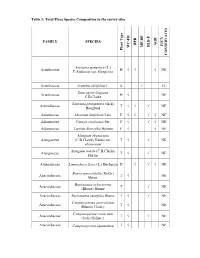
Table 1: Total Flora Species Composition in the Survey Sites
Table 1: Total Flora Species Composition in the survey sites FAMILY SPECIES RPR AGR IUCN FLD-P MF-BF WC-FR Plant Type CONSERVATIO Asystasia gangetica (L.) Acanthaceae H √ √ √ NE T.Anderson ssp. Gangetica Acanthaceae Acanthus ilicifolius L. S √ LC Staurogyne kingiana Acanthaceae H √ NE C.B.Clarke Saurauia pentapetala (Jack) Actinidiaceae T √ √ √ NE Hoogland Adiantaceae Adiantum latifolium Lam. F √ √ √ √ NE Adiantaceae Vittaria ensiformis Sw. F √ √ √ NE Adiantaceae Taenitis dimorpha Holtum F √ √ NE Alangium ebenaceum Alangiaceae (C.B.Clarke) Harms var. T √ √ NE ebenaceum Alangium nobile (C.B.Clarke) Alangiaceae T √ √ NE Harms Alismataceae Limnocharis flava (L.) Buchenau H √ √ √ NE Bouea oppositifolia (Roxb.) Anacardiaceae T √ NE Meisn. Buchanania arborescens Anacardiaceae T √ NE (Blume) Blume Anacardiaceae Buchanania sessifolia Blume T √ √ NE Campnosperma auriculatum Anacardiaceae T √ NE (Blume) Hook.f. Campnosperma coriaceum Anacardiaceae T √ √ NE (Jack) Hallier f. Anacardiaceae Campnosperma squamatum T √ NE FAMILY SPECIES RPR AGR IUCN FLD-P MF-BF WC-FR Plant Type CONSERVATIO Ridl. Dracontomelon dao (Blanco) Anacardiaceae T √ √ NE Merr. & Rolfe Drimycarpus luridus (Hookf) Anacardiaceae T √ NE Ding Hou Gluta aptera (King) Ding Anacardiaceae T √ NE Hou Anacardiaceae Gluta elegans (Wall.) Hook.f. T √ √ NE Anacardiaceae Mangifera gracilipes Hook.f. T √ NE Mangifera macrocarpa Anacardiaceae T √ NE Blume Mangifera magnifica Anacardiaceae T √ NE Kochummen Anacardiaceae Mangifera quadrifida Jack T √ √ NE Melanochyla angustifolia Anacardiaceae T √ NE Hook.f. Melanochyla auriculata Anacardiaceae T √ NE Hook.f. Anacardiaceae Melanochyla bracteata King T √ NE Melanochyla caesia (Blume) Anacardiaceae T √ NE Ding Hou Melanochyla fulvinervis Anacardiaceae T √ NE (Blume) Ding Hou Melanochyla tomentosa Anacardiaceae T √ NE Hook.f. Anacardiaceae Parishia paucijuga Engl.