(Bromus Tectorum) from the Landsat Archive MARK ⁎ Peter J
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Invasive Weeds of the Appalachian Region
$10 $10 PB1785 PB1785 Invasive Weeds Invasive Weeds of the of the Appalachian Appalachian Region Region i TABLE OF CONTENTS Acknowledgments……………………………………...i How to use this guide…………………………………ii IPM decision aid………………………………………..1 Invasive weeds Grasses …………………………………………..5 Broadleaves…………………………………….18 Vines………………………………………………35 Shrubs/trees……………………………………48 Parasitic plants………………………………..70 Herbicide chart………………………………………….72 Bibliography……………………………………………..73 Index………………………………………………………..76 AUTHORS Rebecca M. Koepke-Hill, Extension Assistant, The University of Tennessee Gregory R. Armel, Assistant Professor, Extension Specialist for Invasive Weeds, The University of Tennessee Robert J. Richardson, Assistant Professor and Extension Weed Specialist, North Caro- lina State University G. Neil Rhodes, Jr., Professor and Extension Weed Specialist, The University of Ten- nessee ACKNOWLEDGEMENTS The authors would like to thank all the individuals and organizations who have contributed their time, advice, financial support, and photos to the crea- tion of this guide. We would like to specifically thank the USDA, CSREES, and The Southern Region IPM Center for their extensive support of this pro- ject. COVER PHOTO CREDITS ii 1. Wavyleaf basketgrass - Geoffery Mason 2. Bamboo - Shawn Askew 3. Giant hogweed - Antonio DiTommaso 4. Japanese barberry - Leslie Merhoff 5. Mimosa - Becky Koepke-Hill 6. Periwinkle - Dan Tenaglia 7. Porcelainberry - Randy Prostak 8. Cogongrass - James Miller 9. Kudzu - Shawn Askew Photo credit note: Numbers in parenthesis following photo captions refer to the num- bered photographer list on the back cover. HOW TO USE THIS GUIDE Tabs: Blank tabs can be found at the top of each page. These can be custom- ized with pen or marker to best suit your method of organization. Examples: Infestation present On bordering land No concern Uncontrolled Treatment initiated Controlled Large infestation Medium infestation Small infestation Control Methods: Each mechanical control method is represented by an icon. -

Oregon City Nuisance Plant List
Nuisance Plant List City of Oregon City 320 Warner Milne Road , P.O. Box 3040, Oregon City, OR 97045 Phone: (503) 657-0891, Fax: (503) 657-7892 Scientific Name Common Name Acer platanoides Norway Maple Acroptilon repens Russian knapweed Aegopodium podagraria and variegated varieties Goutweed Agropyron repens Quack grass Ailanthus altissima Tree-of-heaven Alliaria officinalis Garlic Mustard Alopecuris pratensis Meadow foxtail Anthoxanthum odoratum Sweet vernalgrass Arctium minus Common burdock Arrhenatherum elatius Tall oatgrass Bambusa sp. Bamboo Betula pendula lacinata Cutleaf birch Brachypodium sylvaticum False brome Bromus diandrus Ripgut Bromus hordeaceus Soft brome Bromus inermis Smooth brome-grasses Bromus japonicus Japanese brome-grass Bromus sterilis Poverty grass Bromus tectorum Cheatgrass Buddleia davidii (except cultivars and varieties) Butterfly bush Callitriche stagnalis Pond water starwort Cardaria draba Hoary cress Carduus acanthoides Plumeless thistle Carduus nutans Musk thistle Carduus pycnocephalus Italian thistle Carduus tenufolius Slender flowered thistle Centaurea biebersteinii Spotted knapweed Centaurea diffusa Diffuse knapweed Centaurea jacea Brown knapweed Centaurea pratensis Meadow knapweed Chelidonium majou Lesser Celandine Chicorum intybus Chicory Chondrilla juncea Rush skeletonweed Cirsium arvense Canada Thistle Cirsium vulgare Common Thistle Clematis ligusticifolia Western Clematis Clematis vitalba Traveler’s Joy Conium maculatum Poison-hemlock Convolvulus arvensis Field Morning-glory 1 Nuisance Plant List -

Phylogenetic Analyses Reveal the Shady History of C4 Grasses Erika J
Phylogenetic analyses reveal the shady history of C4 grasses Erika J. Edwardsa,1 and Stephen A. Smithb aDepartment of Ecology and Evolutionary Biology, Brown University, Providence, RI 02912; and bNational Evolutionary Synthesis Center, Durham, NC 27705 Edited by Michael J. Donoghue, Yale University, New Haven, CT, and approved December 31, 2009 (received for review August 24, 2009) Grasslands cover more than 20% of the Earth's terrestrial surface, has provided a strong selection pressure for C4 evolution in and their rise to dominance is one of the most dramatic events of eudicots (4). Grasses have long been viewed as an interesting biome evolution in Earth history. Grasses possess two main photo- exception to this pattern (9). Significant positive correlations synthetic pathways: the C3 pathway that is typical of most plants between C4 grass abundance and growing season temperature and a specialized C4 pathway that minimizes photorespiration and have been documented at both continental and regional scales thus increases photosynthetic performance in high-temperature (10–13); C4 grasses dominate tropical grasslands and savannas and/or low-CO2 environments. C4 grasses dominate tropical and but are virtually absent from cool-temperate grasslands and subtropical grasslands and savannas, and C3 grasses dominate the steppes. Furthermore, both experimental measurements of world's cooler temperate grassland regions. This striking pattern photosynthetic light use efficiency (termed “quantum yield”), has been attributed to C4 physiology, with the implication that the and predictions of leaf models of C3 and C4 photosynthesis evolution of the pathway enabled C4 grasses to persist in warmer provide strong evidence that C4 grasses outperform C3 grasses at climates than their C3 relatives. -

1 Supplementary Information for Invasive Grasses Increase
Supplementary Information For Invasive grasses increase fire occurrence and frequency across U.S. ecoregions Emily J. Fusco1*, John T. Finn2, Jennifer K. Balch3,4, R. Chelsea Nagy3, Bethany A. Bradley1,2 Affiliations: 1 Graduate Program in Organismic and Evolutionary Biology, University of Massachusetts- Amherst, Amherst, Massachusetts, 01003, USA 2 Department of Environmental Conservation, University of Massachusetts- Amherst, Amherst, Massachusetts, 01003, USA 3 Earth Lab, University of Colorado- Boulder, Boulder, Colorado, 80309, USA 4 Department of Geography, University of Colorado-Boulder, Boulder, Colorado, 80309, USA Correspondence to: [email protected] This PDF file includes: Figure S1 Tables S1 to S4 SI References 1 www.pnas.org/cgi/doi/10.1073/pnas.1908253116 Supplemental Table S1: A list of 176 non-native invasive grass and other graminoid species as listed by the Invasive Plant Atlas of the United States (1). For each species, we conducted a Web of Science (WOS) search and recorded whether there was literature suggesting the species altered fire regimes (Yes/No). For each fire promoting species in WOS, we supplemented our determination of whether that species was a fire promoter using the Fire Effects Information System (FEIS; 2). For each species designated as a fire promoter, we searched for available spatial data, and kept only species that were both fire-promoting with spatial data for our analysis. Final species used are highlighted in yellow. WOS FEIS Fire Data Keep for Scientific Name Common Name(s) Search Database Promoter Available Analysis Achnatherum punagrass No - No - No brachychaetum Godr. Barkworth Aegilops cylindrica Host jointed goatgrass No - No - No Aegilops ovate goatgrass No - No - No geniculata Roth Aegilops triuncialis L. -

And Mission Blue Butterfly Populations Found at Milagra Ridge and the Mission Blue Butterfly Population at Marin Headlands Are Managed by the GGNRA
San Bruno Elfin Butterfly (Callophrys mossii bayensis) and Mission Blue Butterfly (Icaricia icarioides missionensis) 5-Year Review: Summary and Evaluation Photo by Patrick Kobernus: Adult male mission blue butterfly. Sacramento Fish and Wildlife Field Office U.S. Fish and Wildlife Service Sacramento, California February 2010 5-YEAR REVIEW San Bruno Elfin Butterfly (Callophrys mossii bayensis) and Mission blue butterfly (Icaricia icarioides missionensis) I. GENERAL INFORMATION Purpose of 5-Year Reviews: The U.S. Fish and Wildlife Service (Service) is required by section 4(c)(2) of the Endangered Species Act (Act) to conduct a status review of each listed species at least once every 5 years. The purpose of a 5-year review is to evaluate whether or not the species’ status has changed since it was listed (or since the most recent 5-year review). Based on the 5-year review, we recommend whether the species should be removed from the list of endangered and threatened species, be changed in status from endangered to threatened, or be changed in status from threatened to endangered. Our original listing of a species as endangered or threatened is based on the existence of threats attributable to one or more of the five threat factors described in section 4(a)(1) of the Act, and we must consider these same five factors in any subsequent consideration of reclassification or delisting of a species. In the 5-year review, we consider the best available scientific and commercial data on the species, and focus on new information available since the species was listed or last reviewed. -

Taxonomic Revision of Blumeria Based on Multi-Gene DNA Sequences, Host Preferences and Morphology
Mycoscience: Advance Publication doi: 10.47371/mycosci.2020.12.003 Full Paper (Received July 3, 2020; Accepted December 9, 2020) J-STAGE Advance Published Date: March 13, 2021 Full paper Taxonomic revision of Blumeria based on multi-gene DNA sequences, host preferences and morphology Miao Liua,d,*, Uwe Braunb,d, Susumu Takamatsuc, Sarah Hambletona, Parivash Shoukouhia, Kassandra R. Bissona, Keith Hubbarda a Biodiversity and Bioresources, Ottawa Research and Development Centre, Agriculture and Agri-Food Canada, Ottawa, Ontario K1A 0C6, Canada. b Martin Luther University, Institute of Biology, Department of Geobotany, Herbarium, Halle (Saale) 06099, Germany. c Faculty of Bioresources, Mie University, Tsu 514-8507, Japan. d First two authors should be considered as havingPublication equal contribution. *Corresponding author email: [email protected] Text: 61 pages; tables: 1; figures: 9 Supplemental materials: 4 Supplementary Figures, 1 Supplementary Appendix text Advance - 1 - Mycoscience: Advance Publication ABSTRACT A taxonomic revision of the hitherto monotypic genus Blumeria was conducted incorporating multi-gene sequence analyses, host preference data and morphological criteria. The sequenced loci included rDNA ITS, partial chitin synthase gene (CHS1), as well as fragments of two unnamed orthologous genes (Bgt-1929, Bgt-4572). The combined evidence led to a reassessment and a new neotypification of B. graminis s. str. (emend.), and the description of seven additional species, viz. B. americana sp. nov. (mainly on hosts of the Triticeae), B. avenae sp. nov. (on Avena spp.), B. bromi-cathartici sp. nov. (on Bromus catharticus), B. bulbigera comb. nov. (on Bromus spp.), B. dactylidis sp. nov. (on Dactylis glomerata as the main host, but also on various other hosts), B. -

First Record of Eriochloa Villosa (Thunb.) Kunth in Austria and Notes on Its Distribution and Agricultural Impact in Central Europe
BioInvasions Records (2020) Volume 9, Issue 1: 8–16 CORRECTED PROOF Research Article First record of Eriochloa villosa (Thunb.) Kunth in Austria and notes on its distribution and agricultural impact in Central Europe Swen Follak1,*, Michael Schwarz2 and Franz Essl3 1Institute for Sustainable Plant Production, Austrian Agency for Health and Food Safety, Vienna, Austria 2Data, Statistics and Risk Assessment, Austrian Agency for Health and Food Safety, Vienna, Austria 3Division of Conservation Biology, Vegetation and Landscape Ecology, University of Vienna, Vienna, Austria Author e-mails: [email protected] (SF), [email protected] (MS), [email protected] (FE) *Corresponding author Citation: Follak S, Schwarz M, Essl F (2020) First record of Eriochloa villosa Abstract (Thunb.) Kunth in Austria and notes on its distribution and agricultural impact in Eriochloa villosa is native to temperate Eastern Asia and is an emerging weed in Central Europe. BioInvasions Records 9(1): Central Europe. Its current distribution in Central Europe was analyzed using 8–16, https://doi.org/10.3391/bir.2020.9.1.02 distribution data from the literature and data collected during field trips. In 2019, E. Received: 6 September 2019 villosa was recorded for the first time in Austria. It was found in a crop field in Accepted: 28 November 2019 Unterretzbach in Lower Austria (Eastern Austria). So far, the abundance of E. villosa in the weed communities in Austria and the neighboring Czech Republic is low and Published: 21 February 2020 thus, its present agricultural impact can be considered limited. However, in Romania Handling editor: Quentin Groom and Hungary, the number of records of E. -

Table of Contents
Aliso: A Journal of Systematic and Evolutionary Botany Volume 23 | Issue 1 Article 2 2007 Table of Contents Follow this and additional works at: http://scholarship.claremont.edu/aliso Recommended Citation (2007) "Table of Contents," Aliso: A Journal of Systematic and Evolutionary Botany: Vol. 23: Iss. 1, Article 2. Available at: http://scholarship.claremont.edu/aliso/vol23/iss1/2 PREFACE vii BROMELIACEAE 1 Phylogeny, adaptive radiation, and historical biogeography of Bromeliaceae inferred from ndhF 3 sequence data Thomas J. Givnish, Kendra C. Millam, Paul E. Berry, and Kenneth J. Sytsma Systematics of Bromelioideae (Bromeliaceae)—evidence from molecular and anatomical studies 27 Ralf Horres, Katharina Schulte, Kurt Weising, and Georg Zizka The jeweled armor of Tillandsia—multifaceted or elongated trichomes provide photoprotection 44 Simon Pierce CYPERACEAE, JUNCACEAE, AND THURNIACEAE 53 The Juncaceae-Cyperaceae interface: a combined plastid sequence analysis 55 Eleanor Jones, David A. Simpson, Trevor R. Hodkinson, Mark W. Chase, and John A. N. Parnell Chromosome evolution in Cyperales 62 Eric H. Roalson, Andrew G. McCubbin, and Richard Whitkus Phylogeny of Cyperaceae based on DNA sequence data—a new rbcL analysis 72 David A. Simpson, A. Muthama Muasya, Marccus V. Alves, Jeremy J. Bruhl, Sandra Dhooge, Mark W. Chase, Carol A. Furness, Kioumars Ghamkhar, Paul Goetghebeur, Trevor R. Hodkinson, Adam D. Marchant, Anton A. Reznicek, Roland Nieuwborg, Eric H. Roalson, Erik Smets, Julian R. Starr, William W. Thomas, Karen L. Wilson, and Xiufu Zhang Phylogenetic implications of a unique 5.8S nrDNA insertion in Cyperaceae 84 Julian R. Starr, Genevie`ve Gravel, Anne Bruneau, and A. Muthama Muasya Towards a comprehensive survey of C3 and C4 photosynthetic pathways in Cyperaceae 99 Jeremy J. -

Diversity and Management of Russian-Thistle (Salsola Tragus L.) in the Dryland Cropping Systems of the Inland Pacific Northwest
DIVERSITY AND MANAGEMENT OF RUSSIAN-THISTLE (SALSOLA TRAGUS L.) IN THE DRYLAND CROPPING SYSTEMS OF THE INLAND PACIFIC NORTHWEST By JOHN FORREST SPRING A dissertation submitted in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY WASHINGTON STATE UNIVERSITY Department of Crop and Soil Science MAY 2017 © Copyright by JOHN FORREST SPRING, 2017 All Rights Reserved © Copyright by JOHN FORREST SPRING, 2017 All Rights Reserved To the Faculty of Washington State University: The members of the Committee appointed to examine the dissertation of JOHN FORREST SPRING find it satisfactory and recommend that it be accepted. _________________________________________ Drew J. Lyon, Ph.D., Chair _________________________________________ Ian C. Burke, Ph.D. _________________________________________ Eric H. Roalson, Ph.D. _________________________________________ Frank L. Young, Ph.D. ii ACKNOWLEDGEMENTS Thank you to all that contributed to the conduct of this work, and to the education of a would-be scientist: my advisor, Drew Lyon, and committee members, program technicians, professors, and fellow graduate students. iii DIVERSITY AND MANAGEMENT OF RUSSIAN-THISTLE (SALSOLA TRAGUS L.) IN THE DRYLAND CROPPING SYSTEMS OF THE INLAND PACIFIC NORTHWEST Abstract by John Forrest Spring, Ph.D. Washington State University May 2017 Chair: Drew J. Lyon Russian-thistle (Salsola tragus L.) is one of the most troublesome weed species in the low- and intermediate-precipitation dryland wheat-fallow cropping zones of the inland Pacific Northwest (PNW). High levels of morphological diversity typify the species on global, continental and regional scales. Previous research in California found this variability to encompass a largely cryptic complex of five distinct species in populations of Salsola in that state. -

(Poaceae) and Characterization
EVOLUTION AND DEVELOPMENT OF VEGETATIVE ARCHITECTURE: BROAD SCALE PATTERNS OF BRANCHING ACROSS THE GRASS FAMILY (POACEAE) AND CHARACTERIZATION OF ARCHITECTURAL DEVELOPMENT IN SETARIA VIRIDIS L. P. BEAUV. By MICHAEL P. MALAHY Bachelor of Science in Biology University of Central Oklahoma Edmond, Oklahoma 2006 Submitted to the Faculty of the Graduate College of the Oklahoma State University in partial fulfillment of the requirements for the Degree of MASTER OF SCIENCE July, 2012 EVOLUTION AND DEVELOPMENT OF VEGETATIVE ARCHITECTURE: BROAD SCALE PATTERNS OF BRANCHING ACROSS THE GRASS FAMILY (POACEAE) AND CHARACTERIZATION OF ARCHITECTURAL DEVELOPMENT IN WEEDY GREEN MILLET ( SETARIA VIRIDIS L. P. BEAUV.) Thesis Approved: Dr. Andrew Doust Thesis Adviser Dr. Mark Fishbein Dr. Linda Watson Dr. Sheryl A. Tucker Dean of the Graduate College I TABLE OF CONTENTS Chapter Page I. Evolutionary survey of vegetative branching across the grass family (poaceae) ... 1 Introduction ................................................................................................................... 1 Plant Architecture ........................................................................................................ 2 Vascular Plant Morphology ......................................................................................... 3 Grass Morphology ....................................................................................................... 4 Methods ....................................................................................................................... -

Ecologically-Based Manipulation Practices for Managing Bromus Tectorum-Infested Rangelands
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 8-2011 Ecologically-Based Manipulation Practices for Managing Bromus tectorum-infested Rangelands Beth Fowers Utah State University Follow this and additional works at: https://digitalcommons.usu.edu/etd Part of the Environmental Sciences Commons Recommended Citation Fowers, Beth, "Ecologically-Based Manipulation Practices for Managing Bromus tectorum-infested Rangelands" (2011). All Graduate Theses and Dissertations. 1019. https://digitalcommons.usu.edu/etd/1019 This Thesis is brought to you for free and open access by the Graduate Studies at DigitalCommons@USU. It has been accepted for inclusion in All Graduate Theses and Dissertations by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. ECOLOGICALLY-BASED MANIPULATION PRACTICES FOR MANAGING BROMUS TECTORUM-INFESTED RANGELANDS by Beth Fowers A thesis submitted in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in Range Science Approved: ________________________ ________________________ Dr. Christopher A. Call Dr. Thomas A. Monaco Major Professor Committee Member ________________________ ________________________ Dr. Corey Ransom Dr. Mark R. McLellan Committee Member Vice President for Research and Dean of the School of Graduate Studies UTAH STATE UNIVERSITY Logan, Utah 2011 ii Copyright © Beth Fowers 2011 All Rights Reserved iii ABSTRACT Ecologically-Based Manipulation Practices for Managing Bromus tectorum-infested Rangelands by Beth Fowers, Master of Science Utah State University, 2011 Major Professor: Christopher A. Call Department: Wildland Resources Cheatgrass (Bromus tectorum) is an invasive annual grass common in several semiarid plant communities in the western U.S. B. tectorum presence increases fire frequency and size, reducing species diversity, and leading to annual species-dominated systems with inconsistent livestock forage potential and degraded wildlife habitat value. -
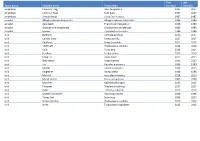
Taxon Group Common Name Taxon Name First Recorded Last
First Last Taxon group Common name Taxon name recorded recorded amphibian Common Frog Rana temporaria 1987 2017 amphibian Common Toad Bufo bufo 1987 2017 amphibian Smooth Newt Lissotriton vulgaris 1987 1987 annelid Alboglossiphonia heteroclita Alboglossiphonia heteroclita 1986 1986 annelid duck leech Theromyzon tessulatum 1986 1986 annelid Glossiphonia complanata Glossiphonia complanata 1986 1986 annelid leeches Erpobdella octoculata 1986 1986 bird Bullfinch Pyrrhula pyrrhula 2016 2017 bird Carrion Crow Corvus corone 2017 2017 bird Chaffinch Fringilla coelebs 2015 2017 bird Chiffchaff Phylloscopus collybita 2014 2016 bird Coot Fulica atra 2014 2014 bird Fieldfare Turdus pilaris 2015 2015 bird Great Tit Parus major 2015 2015 bird Grey Heron Ardea cinerea 2013 2017 bird Jay Garrulus glandarius 1999 1999 bird Kestrel Falco tinnunculus 1999 2015 bird Kingfisher Alcedo atthis 1986 1986 bird Mallard Anas platyrhynchos 2014 2015 bird Marsh Harrier Circus aeruginosus 2000 2000 bird Moorhen Gallinula chloropus 2015 2015 bird Pheasant Phasianus colchicus 2017 2017 bird Robin Erithacus rubecula 2017 2017 bird Spotted Flycatcher Muscicapa striata 1986 1986 bird Tawny Owl Strix aluco 2006 2015 bird Willow Warbler Phylloscopus trochilus 2015 2015 bird Wren Troglodytes troglodytes 2015 2015 bird Yellowhammer Emberiza citrinella 2000 2000 conifer Douglas Fir Pseudotsuga menziesii 2004 2004 conifer European Larch Larix decidua 2004 2004 conifer Lawson's Cypress Chamaecyparis lawsoniana 2004 2004 conifer Scots Pine Pinus sylvestris 1986 2004 crustacean