The Biogeomorphological Life Cycle of Poplars During the Fluvial Biogeomorphological Succession: a Special Focus on Populus Nigra L
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Assessment of Forest Pests and Diseases in Protected Areas of Georgia Final Report
Assessment of Forest Pests and Diseases in Protected Areas of Georgia Final report Dr. Iryna Matsiakh Tbilisi 2014 This publication has been produced with the assistance of the European Union. The content, findings, interpretations, and conclusions of this publication are the sole responsibility of the FLEG II (ENPI East) Programme Team (www.enpi-fleg.org) and can in no way be taken to reflect the views of the European Union. The views expressed do not necessarily reflect those of the Implementing Organizations. CONTENTS LIST OF TABLES AND FIGURES ............................................................................................................................. 3 ABBREVIATIONS AND ACRONYMS ...................................................................................................................... 6 EXECUTIVE SUMMARY .............................................................................................................................................. 7 Background information ...................................................................................................................................... 7 Literature review ...................................................................................................................................................... 7 Methodology ................................................................................................................................................................. 8 Results and Discussion .......................................................................................................................................... -

Zoogeomorphology & Ecosystem Engineering
Zoogeomorphology & Ecosystem Engineering The 42nd Annual Binghamton Geomorphology Symposium October 21 – 23, 2011 University of South Alabama, Mobile, Alabama National Science University of Texas State University- Foundation South Alabama San Marcos Zoogeomorphology and Ecosystem Engineering The 42nd Annual Binghamton Geomorphology Symposium University of South Alabama, Mobile, Alabama October 21 – 23, 2011 Conference Organizers: David R. Butler Department of Geography, Texas State University-San Marcos Carol F. Sawyer Department of Earth Sciences, University of South Alabama Conference Sponsors National Science Foundation, GEO/EAR, Geomorphology & Land-Use Dynamics Department of Earth Sciences, University of South Alabama College of Arts and Sciences, University of South Alabama Department of Geography, Texas State University-San Marcos Field Trip Assistance Dr. Mimi Fearn, Ms. Sherall Cornwell, Ms. Karen Jordan, Ms. Tela O’Rourke, and Mr. Donald Brinkman, Department of Earth Sciences, University of South Alabama We wish to thank the following people for their assistance in organizing this symposium: Catherine Drake and the Center for Continuing Education & Conference Services staff, Univ. South Alabama Department of Earth Sciences faculty, staff, and students, including Dr. Mimi Fearn, Dr. Jim Connors, Sherall Cornwell, Chris Reese, Donald Brinkman, Tela O’Rourke, and Reva Stienstraw-Hitchcock Dr. Phil Suckling, Chair, Department of Geography, Texas State University-San Marcos Dr. Jack Vitek, Texas A&M University Dr. Richard Marston, -

Biomechanical and Biochemical Effects Recorded in the Tree Root Zone – Soil Memory, Historical Contingency and Soil Evolution Under Trees
Plant Soil (2018) 426:109–134 https://doi.org/10.1007/s11104-018-3622-9 REGULAR ARTICLE Biomechanical and biochemical effects recorded in the tree root zone – soil memory, historical contingency and soil evolution under trees Łukasz Pawlik & Pavel Šamonil Received: 17 September 2017 /Accepted: 1 March 2018 /Published online: 15 March 2018 # The Author(s) 2018 Abstract increase in soil spatial complexity. We hypothesized that Background and aims The changing soils is a never- trees can be a strong local factor intensifying, blocking ending process moderated by numerous biotic and abi- or modifying pedogenetic processes, leading to local otic factors. Among these factors, trees may play a changes in soil complexity (convergence, divergence, critical role in forested landscapes by having a large or polygenesis). These changes are hypothetically con- imprint on soil texture and chemical properties. During trolled by regionally predominating soil formation their evolution, soils can follow convergent or divergent processes. development pathways, leading to a decrease or an Methods To test the main hypothesis, we described the pedomorphological features of soils under tree stumps of fir, beech and hemlock in three soil regions: Haplic Highlights Cambisols (Turbacz Reserve, Poland), Entic Podzols 1) The architecture of tree root systems controls soil physical and (Žofínský Prales Reserve, Czech Republic) and Albic chemical properties. Podzols (Upper Peninsula, Michigan, USA). Soil pro- 2) The predominating pedogenetic process significantly modifies files under the stumps, as well as control profiles on sites the effect of trees on soil. 3) Trees are a factor in polygenesis in Haplic Cambisols at the currently not occupied by trees, were analyzed in the pedon scale. -
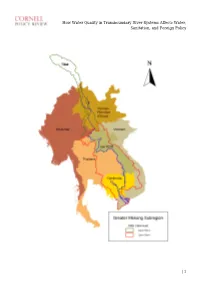
How Water Quality in Transboundary River Systems Affects Water, Sanitation, and Foreign Policy
How Water Quality in Transboundary River Systems Affects Water, Sanitation, and Foreign Policy | 1 How Water Quality in Transboundary River Systems Affects Water, Sanitation, and Foreign Policy David Tipping, 2001 By David C. Tipping Edited by Yeareen Yun Disclaimer: The views and opinions expressed in this article are those of the author and do not necessarily reflect the official policy or position of any agency of the Australian government. Assumptions made within the analysis are not reflective of the position of any Australian government entity, or other organization or professional association. 1. INTRODUCTION Access to adequate water supply and sanitation is the core premise of local level water security. Effective management of transboundary river basin systems and water quality risks is therefore fundamental to social progress and quality of life. Improved water quality management benefits many individual lives in riparian nations, and, as demonstrated by the annual new year blessing of the fish migrations, society at large throughout the Mekong River Basin. In 2001, the author investigated the use of sustainable development indicators to improve the institutional effectiveness of international environmental management regimes. A new framework was designed to evaluate beneficial uses of water. In addition, a case study was developed on the Lower Mekong River Basin system, which integrated measures of water and environmental quality and socio-economic development. The research objectives were: (1) improving the understanding of water quality issues; (2) benchmarking water resources management performance at local, national and regional levels; and (3) enhancing technical and administrative capabilities of transboundary river basin management regimes through capacity development focused on the achievement of sustainable development objectives, and obligations and duties under international law. -

Assessment of Forest Pests and Diseases in Native Boxwood Forests of Georgia Final Report
Assessment of Forest Pests and Diseases in Native Boxwood Forests of Georgia Final report Dr. Iryna Matsiakh Forestry Department, Ukrainian National Forestry University (Lviv) Tbilisi 2016 TABLE OF CONTENT LIST OF TABLES AND FIGURES .................................................................................................................................. 2 ABBREVIATIONS AND ACRONYMS ........................................................................................................................... 5 EXECUTIVE SUMMARY .................................................................................................................................................. 6 INTRODUCTION .............................................................................................................................................................. 10 1. BACKGROUND INFORMATION ............................................................................................................................ 11 1.1. Biodiversity of Georgia ........................................................................................................................................ 11 1.2. Forest Ecosystems .................................................................................................................................................. 12 1.3. Boxwood Forests in Forests Habitat Classification ................................................................................. 14 1.4. Georgian Forests Habitat in the Context of Climate Change -

Guide 3 – Fish Farmer's Guide to Combating Parasitic
GUIDE 3 – FISH FARMER’S GUIDE TO COMBATING PARASITIC INFECTIONS IN COMMON CARP AQUACULTURE e-NIPO: 833-20-103-X A Series of ParaFishControl Guides to Combating Fish Parasite Infections in Aquaculture. Guide 3 This project has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 634429 (ParaFishControl). This output reflects only the author’s view and the European Union cannot be held responsible for any use that may be made of the information contained therein. Wherever the fish are, that's where we go. “ Richard Wagner “ Common carp is the third most cultivated freshwater species in the world. Carp aquaculture is usually performed in a semi-intensive manner, in earthen ponds, where parasitic diseases can easily compromise fish health, especially in the hot summer months, leading to production and economic losses. This guide provides useful information about the biological background of five parasites, their diagnostics and control measures. © A.S. Holzer List of Authors Dr Astrid S. Holzer, Principal Investigator and Team Leader Institute of Parasitology Biology Centre of the Czech Academy of Sciences, Czech Republic Email: [email protected] Dr Pavla Bartošová-Sojková, Researcher Institute of Parasitology Biology Centre of the Czech Academy of Sciences, Czech Republic Email: [email protected] Honorary Prof. Csaba Székely, Scientific Advisor and Team Leader Institute for Veterinary Medical Research, Centre for Agricultural Research, (former Hungarian Academy of Sciences), Hungary Email: [email protected] Dr Gábor Cech, Senior Researcher, Institute for Veterinary Medical Research, Centre for Agricultural Research, (former Hungarian Academy of Sciences), Hungary Email: [email protected] Dr Kálmán Molnár, Retired Scientific Advisor, Fish Pathology and Parasitology Research Team, Institute for Veterinary Medical Research, Centre for Agricultural Research (former Hungarian Academy of Sciences), Hungary Prof. -

Difference Between Haplontic and Diplontic Life Cycles
Difference Between Haplontic and Diplontic Life Cycles www.differencebetween.com Key Difference – Haplontic vs Diplontic Life Cycles In the context of biology, a biological life cycle is a sequence of changes a particular organism undergoes through means of reproduction (sexual or asexual) which finally returns to the original starting phase. This procedure differs from one organism to the other. During sexual reproduction, the life cycle includes the change of ploidy; the alternation of haploid (n) and diploid (2n) stages. Meiosis occurs during the change over from a diploid stage to a haploid stage. With regards to change of ploidy, life cycles are of three types. They are, haplontic, diplontic and haplodiplontic. In a haplontic life cycle, the haploid stage is typically multicellular and results in the formation of a diploid (2n) cell, which is a zygote. The zygote undergoes meiosis, which results in the formation of haploid (n) cells. In a diplontic life cycle, the diploid stage is typically multicellular, and meiosis occurs during gamete formation which results in the production of haploid (n) gametes. During fertilization, the haploid (n) gametes fuse together in the formation of a diploid (2n) zygote, and it mitotically divides and produces a multicellular diploid (2n) organism. This is the key difference between haplontic and diplontic life cycles. What is a Haplontic Life Cycle? Haplontic life cycle involves the formation of a haploid (n) single cell by the meiosis of a diploid (2n) zygote. This phenomenon could be explained with sporic meiosis – the process of formation of spores. In this process, the zygote mitotically divides and produces multicellular sporophyte which is diploid (2n). -

FINAL COMPASS Aquaculture Roundtable Brief
Science in Action: Exploring the Future of U.S. Aquaculture A COMPASS Roundtable on Ocean Aquaculture As the population continues to expand—both domestically and globally—identifying secure, safe sources of protein is a critical need. With two-thirds of the planet covered in water, it is logical to turn to the ocean as an arena for producing food. Globally, aquaculture is the fastest growing food sector,[i] underscoring the importance of understanding the scientific, policy, and social implications of ocean aquaculture. As with all types of cultivated food production, there are complex and interwoven challenges and opportunities in ocean aquaculture.[ii] Indigenous knowledge and current research can answer questions around environmental safeguards, ecological impacts, long-term sustainable use of marine resources, and the social dimensions of ocean aquaculture. While we’ve developed a deeper scientific understanding of aquaculture, there remains a gap between the state of the science, federal policy, and public perceptions of ocean aquaculture in the U.S.[iii] In order to help provide research insights on the science related to aquaculture, COMPASS convened a roundtable discussion with scientists and policymakers in July 2019. The Roundtable examined ways that science can inform safe, sustainable, and socially acceptable ocean aquaculture in the United States. In preparation, COMPASS staff examined the U.S. aquaculture landscape by speaking with more than 50 scientists, managers, policymakers, and tribal representatives. These stage-setting conversations reflected the key concerns surrounding ocean aquaculture such as best management practices, economics, pollution, interactions with wild populations, and climate change. They also highlighted some of the scientific, technological, and cultural advancements in contemporary aquaculture that could address and reduce some of the perceived risks. -

Riparian Vegetation and the Fluvial Environment: a Biogeographic Perspective
Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the Treatise on Geomorphology, the copy attached is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use. This includes without limitation use in instruction at your institution, distribution to specific colleagues, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial Bendix J., and Stella J.C. Riparian Vegetation and the Fluvial Environment: A Biogeographic Perspective. In: John F. Shroder (Editor-in-chief), Butler, D.R., and Hupp, C.R. (Volume Editors). Treatise on Geomorphology, Vol 12, Ecogeomorphology, San Diego: Academic Press; 2013. p. 53-74. © 2013 Elsevier Inc. All rights reserved. 12.5 Riparian Vegetation and the Fluvial Environment: A Biogeographic Perspective J Bendix, Syracuse University, Syracuse, NY, USA JC Stella, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA r 2013 Elsevier Inc. All rights reserved. 12.5.1 Introduction 53 12.5.2 Early History: -

Survivability of Moss and Fungal Spores in Tests Simulating
Biological Sciences in Space, Vol.25 No.2-4, 83-92,Takahashi, 2011 Y. et al. Original Paper Survivability of Moss and Fungal spores irradiated for 30 min. On the other hand, a colony did not develop if the spores were Spores in Tests Simulating taken off the beads and irradiated for 10 min. Conditions of the ISS Outer Wall This indicates that UV does not penetrate to the other side of the beads, and so the spores on Yuichi Takahashi1*, Hirofumi Hashimoto2, that side can be protected from UV radiation. 3 1 ©2011 Jpn. Soc. Biol. Sci. Space; Article ID: Takuo Nakagawa and Shinpei Shibata 112502011 1Department of Astrophysics, Interactive Symbio- sphere Science, Yamagata University Graduate Key words; fungi, moss, spore, survivability, thermal cycle treatment, UV irradiation School of Science and Engineering, Kojirakawa- machi 1-4-12, Yamagata-City, Yamagata 990-8560, Introduction Japan 2 Institute of Space and Astronautical Science, There are many factors that can influence the survivability JAXA, Yoshinodai 3-1-1 Chuo-ku, Sagamihara- of terrestrial life (TL) in interstellar and interplanetary space, City, Kanagawa 252-5210, Japan for example full-spectrum electromagnetic radiation from 3Inspection room, Kojirakawa-Shiseidoh Hospital, the sun, temperature fluctuations, cosmic particle radiation, Higashihara-machi 1-12-26, Yamagata-City, Ya- vacuum, and microgravity. However, space conditions can magata 990-0034, Japan only be partially simulated in ground experiments. Recently, several experiments have investigated the possibility of TL Abstract survival in space, for example EXPOSE-E (ESA Human Space Flight (2010) Expose, http:// smsc.cnes.fr/EXPOSE/), BIORISK (Baranow et al., 2009; Novikova et al., 2010) and To investigate whether terrestrial life (TL) can BIOPAN (De la Torre Noetzel et al., 2007; Devara et al., survive in interstellar and interplanetary space, 2010), while another one, TANPOPO, will take place in the an experiment was performed to simulate near future. -

BIOGEOMORPHOLOGY of COASTAL STRUCTURES: Understanding Interactions Between Hard Substrata and Colonising Organisms As a Tool for Ecological Enhancement
BIOGEOMORPHOLOGY OF COASTAL STRUCTURES: Understanding interactions between hard substrata and colonising organisms as a tool for ecological enhancement Submitted by Martin Andrew Coombes B.Sc. M.Sc. , to the University of Exeter as a thesis for the degree of Doctor of Philosophy in Geography In February 2011 This thesis is available for Library use on the understanding that it is copyright material and that no quotation from the thesis may be published without proper acknowledgement. I certify that all material in this thesis which is not my own work has been identified and that no material has previously been submitted and approved for the award of a degree by this or any other University. Signature: M. A. Coombes --- BLANK --- 2 ABSTRACT Urbanisation is increasingly recognised as a major ecological pressure at the coast. By 2035, the Department for Environment, Food and Rural Affairs will have to spend £1 billion each year on flood defence and erosion control infrastructure if current levels of protection are to be sustained in England and Wales; this represents a substantial commitment to building new hard structures. Ecological research has shown that structures like seawalls, breakwaters, and harbour and port infrastructure are poor surrogates for undisturbed rocky shores. This, alongside substantial international policy drivers, has led to an interest in the ways in which structures might be enhanced for ecological gain. Virtually all of this research has been undertaken by ecologists, while the contribution of geomorphological understanding has not been fully recognised. This thesis presents an assessment of the two-way interactions between colonising organisms and the materials used to build hard coastal structures under a framework of biogeomorphology. -

Biodiversity Effects on Dune and Salt Marsh Biogeomorphology – a Trait-Based Approach
Biodiversity effects on dune and salt marsh biogeomorphology – a trait-based approach Von der Fakultät für Mathematik und Naturwissenschaften der Carl von Ossietzky Universität Oldenburg zur Erlangung des Grades und Titels eines Doktors der Naturwissenschaften (Dr. rer. nat.) angenommene Dissertation von von Julia Bass geboren am 03. Februar 1989 in Bremen Oldenburg 2019 Gutachter Prof. Dr. Michael Kleyer Zweitgutachter Prof. Dr. Gerhard Zotz Tag der Disputation 13.11.2019 Contents Summary .........................................................................................................................1 Zusammenfassung ..........................................................................................................4 1 Introduction ............................................................................................................8 1.1 The concept of biogeomorphology ..................................................................8 1.2 The trait-based perspective in ecology ..........................................................13 1.3 Combining the biogeomorphic succession model and the functional trait approach .........................................................................................................18 1.4 Barrier islands and Halligen in the Wadden Sea ...........................................25 1.5 Research objectives .......................................................................................35 2 Morphological plasticity of dune pioneer plants in response to timing and magnitude