Atlantic Herring (Clupea Harengus) in the Irish and Celtic Seas; Tracing Populations of the Past and Present
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cod Fact Sheet
R FACT SHEET Cod (Gadus morhua) Image taken from (Cohen et al. 1990) Introduction Cod (Gadus morhua) is generally considered a demersal fish although its habitat may become pelagic under certain hydrographic conditions, when feeding or spawning. It is widely distributed throughout the north Atlantic and Arctic regions in a variety of habitats from shoreline to continental shelf, in depths to 600m (Cohen et al. 1990). The Irish Sea stock spawns at two main sites in the western and eastern Irish Sea during February to April (Armstrong et al. 2011). Historically the stock has been commercially important, however in the last decade a decline in SSB and reduced productivity of the stock have led to reduce landings. Reviewed distribution map for Gadus morhua (Atlantic cod), with modelled year 2100 native range map based on IPCC A2 emissions scenario. www.aquamaps.org, version of Aug. 2013. Web. Accessed 28 Jan. 2011 Life history overview Adults are usually found in deeper, colder waters. During the day they form schools and swim about 30-80 m above the bottom, dispersing at night to feed (Cohen et al. 1990; ICES 2005). They are omnivorous; feeding at dawn or dusk on invertebrates and fish, including their own young (Cohen et al. 1990). Adults migrate between spawning, feeding and overwintering areas, mostly within the boundaries of the respective stocks. Large migrations are rare occurrences, although there is evidence for limited seasonal migrations into neighbouring regions, most Irish Sea fish will stay within their management area (ICES 2012). Historical tagging studies indicated spawning site fidelity but with varying degrees of mixing of cod between the Irish Sea, Celtic Sea and west of Scotland/north of Ireland (ICES 2015). -

THE DEVELOPMENT of MESOZOIC SEDIMENTARY IBASINS AROUND the MARGINS of the NORTH ATLANTIC and THEIR HYDROCARBON POTENTIAL D.G. Masson and P.R
INTERNAL DOCUMENT IIL THE DEVELOPMENT OF MESOZOIC SEDIMENTARY IBASINS AROUND THE MARGINS OF THE NORTH ATLANTIC AND THEIR HYDROCARBON POTENTIAL D.G. Masson and P.R. Miles Internal Document No. 195 December 1983 [This document should not be cited in a published bibliography, and is supplied for the use of the recipient only]. iimsTit\jte of \ OCEANOGRAPHIC SCIENCES % INSTITUTE OF OCEANOGRAPHIC SCIENCES Wormley, Godalming, Surrey GU8 5UB (042-879-4141) (Director: Dr. A. S. Laughton, FRS) Bidston Observatory, Crossway, Birkenhead, Taunton, Mersey side L43 7RA Somerset TA1 2DW (051-653-8633) (0823-86211) (Assistant Director: Dr. D. E. Cartwright) (Assistant Director: M.J. Tucker) THE DEVELOPMENT OF MESOZOIC SEDIMENTARY BASINS AROUND THE MARGINS OF THE NORTH ATLANTIC AND THEIR HYDROCARBON POTENTIAL D.G. Masson and P.R. Miles Internal Document No. 195 December 1983 Work carried out under contract to the Department of Energy This document should not be cited in a published bibliography except as 'personal communication', and is supplied for the use of the recipient only. Institute of Oceanographic Sciences, Brook Road, Wormley, Surrey GU8 SUB ABSTRACT The distribution of Mesozoic basins around the margins of the North Atlantic has been summarised, and is discussed in terms of a new earliest Cretaceous palaeogeographic reconstruction for the North Atlantic area. The Late Triassic-Early Jurassic rift basins of Iberia, offshore eastern Canada and the continental shelf of western Europe are seen to be the fragments of a formerly coherent NE trending rift system which probably formed as a result of tensional stress between Europe, Africa and North America. The separation of Europe, North America and Iberia was preceded by a Late Jurassic-Early Cretaceous rifting phase which is clearly distinct from the earlier Mesozoic rifting episode and was little influenced by it. -

Sample Assessment Tasks
SAMPLE ASSESSMENT TASKS MARINE AND MARITIME STUDIES GENERAL YEAR 11 Copyright © School Curriculum and Standards Authority, 2014 This document – apart from any third party copyright material contained in it – may be freely copied, or communicated on an intranet, for non-commercial purposes in educational institutions, provided that the School Curriculum and Standards Authority is acknowledged as the copyright owner, and that the Authority’s moral rights are not infringed. Copying or communication for any other purpose can be done only within the terms of the Copyright Act 1968 or with prior written permission of the School Curriculum and Standards Authority. Copying or communication of any third party copyright material can be done only within the terms of the Copyright Act 1968 or with permission of the copyright owners. Any content in this document that has been derived from the Australian Curriculum may be used under the terms of the Creative Commons Attribution 4.0 International licence. Disclaimer Any resources such as texts, websites and so on that may be referred to in this document are provided as examples of resources that teachers can use to support their learning programs. Their inclusion does not imply that they are mandatory or that they are the only resources relevant to the course. 2014/18740v5 1 Sample assessment task Marine and Maritime Studies – General Year 11 Task 3 — Unit 1 Assessment type: Extended response Conditions Period allowed for completion of the task – three weeks; a combination of in-class and out-of-class time Task weighting: 2.5% of the school mark for this pair of units __________________________________________________________________________________ Marine resource management research (25 marks) Background Commercial fishing is an important contributor to the Western Australian economy. -

The Biology and Ecology of Samson Fish Seriola Hippos
The biology of Samson Fish Seriola hippos with emphasis on the sportfishery in Western Australia. By Andrew Jay Rowland This thesis is presented for the degree of Doctor of Philosophy at Murdoch University 2009 DECLARATION I declare that the information contained in this thesis is the result of my own research unless otherwise cited. ……………………………………………………. Andrew Jay Rowland 2 Abstract This thesis had two overriding aims. The first was to describe the biology of Samson Fish Seriola hippos and therefore extend the knowledge and understanding of the genus Seriola. The second was to uses these data to develop strategies to better manage the fishery and, if appropriate, develop catch-and-release protocols for the S. hippos sportfishery. Trends exhibited by marginal increment analysis in the opaque zones of sectioned S. hippos otoliths, together with an otolith of a recaptured calcein injected fish, demonstrated that these opaque zones represent annual features. Thus, as with some other members of the genus, the number of opaque zones in sectioned otoliths of S. hippos are appropriate for determining age and growth parameters of this species. Seriola hippos displayed similar growth trajectories to other members of the genus. Early growth in S. hippos is rapid with this species reaching minimum legal length for retention (MML) of 600mm TL within the second year of life. After the first 5 years of life growth rates of each sex differ, with females growing faster and reaching a larger size at age than males. Thus, by 10, 15 and 20 years of age, the predicted fork lengths (and weights) for females were 1088 (17 kg), 1221 (24 kg) and 1311 mm (30 kg), respectively, compared with 1035 (15 kg), 1124 (19 kg) and 1167 mm (21 kg), respectively for males. -

Are Signals of Prey Variability Present in the Demographic Characteristics of Crested Tern (Sterna Bergii) Populations in South Australia?
Are signals of prey variability present in the demographic characteristics of crested tern (Sterna bergii) populations in South Australia? Final Report to the South Australian Wildlife Conservation Fund Project No 0256 Authors: L.J. McLeay1,2, *, B. Page1, S.D. Goldsworthy1 and T.M. Ward1 1 South Australian Research and Development Institute, Aquatic Sciences, 2 Hamra avenue West Beach. PO Box 120, Henley Beach, South Australia 5022, Australia. 2 The University of Adelaide, School of Earth and Environmental Sciences, Adelaide, South Australia 5005, Australia. Correspondence to L.J. McLeay: tel: +61 08 8207 5439; fax: +61 08 8207 5406; Email: [email protected] SARDI Aquatic Sciences Publication No. F2008/000964 SARDI Research Report Series No. 312 Disclaimer The authors warrant that they have taken all reasonable care in producing this report. The report has been through the SARDI Aquatic Sciences internal review process, and has been formally approved for release by the Chief Scientist. Although all reasonable efforts have been made to ensure quality, SARDI Aquatic Sciences does not warrant that the information in this report is free from errors or omissions. SARDI Aquatic Sciences does not accept any liability for the contents of this report or for any consequences arising from its use or any reliance placed upon it. © 2008 SARDI AQUATIC SCIENCES This work is copyright. Apart from any use as permitted under the Copyright Act 1968, no part may be reproduced by any process without prior written permission from the author. Authors: L.J. McLeay, B. Page, S.D. Goldsworthy and T.M. Ward Reviewers: Mr P. -

In South Australia – Stock Structure and Adult Movement
SPATIAL MANAGEMENT OF SOUTHERN GARFISH (HYPORHAMPHUS MELANOCHIR) IN SOUTH AUSTRALIA – STOCK STRUCTURE AND ADULT MOVEMENT MA Steer, AJ Fowler, and BM Gillanders (Editors). Final Report for the Fisheries Research and Development Corporation FRDC Project No. 2007/029 SARDI Aquatic Sciences Publication No. F2009/000018-1 SARDI Research Report Series No. 333 ISBN 9781921563089 October 2009 i Title: Spatial management of southern garfish (Hyporhamphus melanochir) in South Australia – stock structure and adult movement Editors: MA Steer, AJ Fowler, and BM Gillanders. South Australian Research and Development Institute SARDI Aquatic Sciences 2 Hamra Avenue West Beach SA 5024 Telephone: (08) 8207 5400 Facsimile: (08) 8207 5406 http://www.sardi.sa.gov.au DISCLAIMER The authors do not warrant that the information in this document is free from errors or omissions. The authors do not accept any form of liability, be it contractual, tortious, or otherwise, for the contents of this document or for any consequences arising from its use or any reliance placed upon it. The information, opinions and advice contained in this document may not relate, or be relevant, to a readers particular circumstances. Opinions expressed by the authors are the individual opinions expressed by those persons and are not necessarily those of the publisher, research provider or the FRDC. © 2009 Fisheries Research and Development Corporation and SARDI Aquatic Sciences. This work is copyright. Apart from any use as permitted under the Copyright Act 1968 (Cwth), no part of this publication may be reproduced by any process, electronic or otherwise, without the specific written permission of the copyright owners. Neither may information be stored electronically in any form whatsoever without such permission. -

Appendices Appendices
APPENDICES APPENDICES APPENDIX 1 – PUBLICATIONS SCIENTIFIC PAPERS Aidoo EN, Ute Mueller U, Hyndes GA, and Ryan Braccini M. 2015. Is a global quantitative KL. 2016. The effects of measurement uncertainty assessment of shark populations warranted? on spatial characterisation of recreational fishing Fisheries, 40: 492–501. catch rates. Fisheries Research 181: 1–13. Braccini M. 2016. Experts have different Andrews KR, Williams AJ, Fernandez-Silva I, perceptions of the management and conservation Newman SJ, Copus JM, Wakefield CB, Randall JE, status of sharks. Annals of Marine Biology and and Bowen BW. 2016. Phylogeny of deepwater Research 3: 1012. snappers (Genus Etelis) reveals a cryptic species pair in the Indo-Pacific and Pleistocene invasion of Braccini M, Aires-da-Silva A, and Taylor I. 2016. the Atlantic. Molecular Phylogenetics and Incorporating movement in the modelling of shark Evolution 100: 361-371. and ray population dynamics: approaches and management implications. Reviews in Fish Biology Bellchambers LM, Gaughan D, Wise B, Jackson G, and Fisheries 26: 13–24. and Fletcher WJ. 2016. Adopting Marine Stewardship Council certification of Western Caputi N, de Lestang S, Reid C, Hesp A, and How J. Australian fisheries at a jurisdictional level: the 2015. Maximum economic yield of the western benefits and challenges. Fisheries Research 183: rock lobster fishery of Western Australia after 609-616. moving from effort to quota control. Marine Policy, 51: 452-464. Bellchambers LM, Fisher EA, Harry AV, and Travaille KL. 2016. Identifying potential risks for Charles A, Westlund L, Bartley DM, Fletcher WJ, Marine Stewardship Council assessment and Garcia S, Govan H, and Sanders J. -
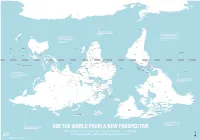
Is Map Is Just As Accurate As the One We're All Used To
ROSS SEA WEDDELL SEA AMUNDSEN SEA ANTARCTICA BELLINGSHAUSEN SEA AMERY ICE SHELF SOUTHERN OCEAN SOUTHERN OCEAN SOUTHERN OCEAN SCOTIA SEA DRAKE PASSAGE FALKLAND ISLANDS Stanley (U.K.) THE RATIO OF LAND TO WATER IN THE SOUTHERN HEMISPHERE BY THE TIME EUROPEANS ADOPTED IS 1 TO 5 THE NORTH-POINTING COMPASS, PTOLEMY WAS A HELLENIC Wellington THEY WERE ALREADY EXPERIENCED NEW TASMAN SEA ZEALAND CARTOGRAPHER WHOSE WORK CHILE IN NAVIGATING WITH REFERENCE TO IN THE SECOND CENTURY A.D. ARGENTINA THE NORTH STAR Canberra Buenos Santiago GREAT AUSTRALIAN BIGHT POPULARIZED NORTH-UP Montevideo Aires SOUTH PACIFIC OCEAN SOUTH ATLANTIC OCEAN URUGUAY ORIENTATION SOUTH AFRICA Maseru LESOTHO SWAZILAND Mbabane Maputo Asunción AUSTRALIA Pretoria Gaborone NEW Windhoek PARAGUAY TONGA Nouméa CALEDONIA BOTSWANA Saint Denis NAMIBIA Nuku’Alofa (FRANCE) MAURITIUS Port Louis Antananarivo MOZAMBIQUE CHANNEL ZIMBABWE Suva Port Vila MADAGASCAR VANUATU MOZAMBIQUE Harare La Paz INDIAN OCEAN Brasília LAKE SOUTH PACIFIC OCEAN FIJI Lusaka TITICACA CORAL SEA GREAT GULF OF BARRIER CARPENTARIA Lilongwe ZAMBIA BOLIVIA FRENCH POLYNESIA Apia REEF LAKE (FRANCE) SAMOA TIMOR SEA COMOROS NYASA ANGOLA Lima Moroni MALAWI Honiara ARAFURA SEA BRAZIL TIMOR LESTE PERU Funafuti SOLOMON Port Dili Luanda ISLANDS Moresby LAKE Dodoma TANGANYIKA TUVALU PAPUA Jakarta SEYCHELLES TANZANIA NEW GUINEA Kinshasa Victoria BURUNDI Bujumbura DEMOCRATIC KIRIBATI Brazzaville Kigali REPUBLIC LAKE OF THE CONGO SÃO TOMÉ Nairobi RWANDA GABON ECUADOR EQUATOR INDONESIA VICTORIA REP. OF AND PRINCIPE KIRIBATI EQUATOR -

Chapter 24. the BAY of BISCAY: the ENCOUNTERING of the OCEAN and the SHELF (18B,E)
Chapter 24. THE BAY OF BISCAY: THE ENCOUNTERING OF THE OCEAN AND THE SHELF (18b,E) ALICIA LAVIN, LUIS VALDES, FRANCISCO SANCHEZ, PABLO ABAUNZA Instituto Español de Oceanografía (IEO) ANDRE FOREST, JEAN BOUCHER, PASCAL LAZURE, ANNE-MARIE JEGOU Institut Français de Recherche pour l’Exploitation de la MER (IFREMER) Contents 1. Introduction 2. Geography of the Bay of Biscay 3. Hydrography 4. Biology of the Pelagic Ecosystem 5. Biology of Fishes and Main Fisheries 6. Changes and risks to the Bay of Biscay Marine Ecosystem 7. Concluding remarks Bibliography 1. Introduction The Bay of Biscay is an arm of the Atlantic Ocean, indenting the coast of W Europe from NW France (Offshore of Brittany) to NW Spain (Galicia). Tradition- ally the southern limit is considered to be Cape Ortegal in NW Spain, but in this contribution we follow the criterion of other authors (i.e. Sánchez and Olaso, 2004) that extends the southern limit up to Cape Finisterre, at 43∞ N latitude, in order to get a more consistent analysis of oceanographic, geomorphological and biological characteristics observed in the bay. The Bay of Biscay forms a fairly regular curve, broken on the French coast by the estuaries of the rivers (i.e. Loire and Gironde). The southeastern shore is straight and sandy whereas the Spanish coast is rugged and its northwest part is characterized by many large V-shaped coastal inlets (rias) (Evans and Prego, 2003). The area has been identified as a unit since Roman times, when it was called Sinus Aquitanicus, Sinus Cantabricus or Cantaber Oceanus. The coast has been inhabited since prehistoric times and nowadays the region supports an important population (Valdés and Lavín, 2002) with various noteworthy commercial and fishing ports (i.e. -

Names of Sub-Areas and Divisions of FAO Fishing Areas 27 and 37 NORTH-EAST ATLANTIC
Names of Sub-areas and Divisions of FAO fishing areas 27 and 37 NORTH-EAST ATLANTIC Subarea I Barents Sea Subarea II Norwegian Sea, Spitzbergen, and Bear Island Division II a Norwegian Sea Division II b Spitzbergen and Bear Island Subarea III Skagerrak, Kattegat, Sound, Belt Sea, and Baltic Sea; the Sound and Belt together known also as the Transition Area Division III a Skagerrak and Kattegat Division III b,c Sound and Belt Sea or Transition Area Division III b (23) Sound Division III c (22) Belt Sea Division III d (24-32) Baltic Sea Subarea IV North Sea Division IV a Northern North Sea Division IV b Central North Sea Division IV c Southern North Sea Subarea V Iceland and Faroes Grounds Division V a Iceland Grounds Division V b Faroes Grounds Subarea VI Rockall, Northwest Coast of Scotland and North Ireland, the Northwest Coast of Scotland and North Ireland also known as the West of Scotland Division VI a Northwest Coast of Scotland and North Ireland or West of Scotland Division VI b Rockall Subarea VII Irish Sea, West of Ireland, Porcupine Bank, Eastern and Western English Channel, Bristol Channel, Celtic Sea North and South, and Southwest of Ireland - East and West Division VII a Irish Sea Division VII b West of Ireland Division VII c Porcupine Bank Division VII d Eastern English Channel Division VII e Western English Channel Division VII f Bristol Channel Division VII g Celtic Sea North Division VII h Celtic Sea South Division VII j South-West of Ireland - East Division VII k South-West of Ireland - West Subarea VIII Bay of Biscay -

Table of Fishes of Sydney Harbour 2019
Table of Fishes of Sydney Harbour 2019 Family Family/Com Species Species Common Notes mon Name Name Acanthuridae Surgeonfishe Acanthurus Eyestripe close s dussumieri Surgeonfish to southern li mit Acanthuridae Acanthurus Orangebloch close to olivaceus Surgeonfish southern limit Acanthuridae Acanthurus Convict close to triostegus Surgeonfish southern limit Acanthuridae Acanthurus Yellowmask xanthopterus Surgeonfish Acanthuridae Paracanthurus Blue Tang not included hepatus in species count Acanthuridae Prionurus Spotted Sawtail maculatus Acanthuridae Prionurus Australian Sawtail microlepidotus Ambassidae Glassfishes Ambassis Port Jackson jacksoniensis glassfish Ambassidae Ambassis marianus Estuary Glassfish Anguillidae Freshwater Anguilla australis Shortfin Eel Eels Anguillidae Anguilla reinhardtii Longfinned Eel Antennariidae Anglerfishes Antennarius Freckled Anglerfish southern limit coccineus Antennariidae Antennarius Giant Anglerfish close to commerson southen limit Antennariidae Antennarius Shaggy Anglerfish southern limit hispidus Antennariidae Antennarius pictus Painted Anglerfish Antennariidae Antennarius striatus Striate Anglerfish Table of Fishes of Sydney Harbour 2019 Antennariidae Histrio histrio Sargassum close to Anglerfish southen limit Antennariidae Porophryne Red-fingered erythrodactylus Anglerfish Aploactinidae Velvetfishes Aploactisoma Southern Velvetfish milesii Aploactinidae Cocotropus Patchwork microps Velvetfish Aploactinidae Paraploactis Bearded Velvetfish trachyderma Aplodactylidae Seacarps Aplodactylus Rock Cale -

1. a Plankton Highway Along the Western Coasts of the UK
HABs themselves should not be used as indicator 1. A plankton highway along of water quality. the western coasts of the UK Main policy implications This work has demonstrated that the stratified regions around the western coasts of the UK and Ireland have particular unique properties and constitute distinct eco-hydrodynamic regions. Such oceanographic regionality is directly relevant to a range of aspects likely to be considered in the Marine Bill, especially Marine Spatial Planning (http://www.defra.gov.uk/corporate/consult/marine bill/index.htm). Use of such knowledge is fundamental to the characterisation (‘typology’) of UK shelf waters (relevant to the proposed European Marine Strategy Directive), and increasing awareness is required of these oceanographic complexities with regard to, for example, the consideration of indicators of ecosystem status (Tett et al., 2004a, b; Larcombe et al. 2004), fisheries management and marine nature conservation. In summer, the density-driven flows at the boundaries of the stratified regions cause a major regional flow, which acts to limit transport between neighbouring eco-hydrodynamic regions. For example, summer transport between the Celtic Sea and the Irish Sea is very limited, while there is strong transport along their boundary (St Georges Channel). There is thus limited potential for transport of plankton across these pathways, Figure 1.1. The combined pathway from the results of three but high potential for transport along them, as field programs, with drifter tracks overlain on the contours of indicated by the occurrence of Karenia mikimotoi bottom density field. along this pathway (Raine, 1993). This mechanism has the potential to transport ‘non- Main science findings indigenous’ species into and around UK waters, into environments that may, in the future, be From early summer (late May) to autumn (mid favourable to their persistence.