Concretions Or Fossils?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Field and Petrographic Study of the Freda
A FIELD AND PETROGRAPHIC STUDY OF THE FREDA FORMATION ALONG THE MONTREAL RIVER, GOGEBIC COUNTY, MICHIGAN fey GORDON R. SMALE A THESIS Submitted in partial fulfillment of the requirements for the degree of Master of Science in Michigan State University East Lansing, Michigan 1958 ProQuest Number: 10008738 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. uest. ProQuest 10008738 Published by ProQuest LLC (2016). Copyright of the Dissertation is held by the Author. All rights reserved. This work is protected against unauthorized copying under Title 17, United States Code Microform Edition © ProQuest LLC. ProQuest LLC. 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, Ml 48106- 1346 A FIELD AND PETROGRAPHIC STUDY OF THE FREDA FORMATION ALONG THE MONTREAL RIVER, GOGEBIC COUNTY, MICHIGAN •"by Gordon R „ Smal'e ABSTRACT The Montreal River provides a geographic boundary between Wisconsin and Michigan on the south shore of Lake Superior. A geologic study, both field and laboratory, was made of the 12,000 foot thickness of steeply dipping Freda for mation exposed along this river. The field study Includes mapping of sedimentary structures In an effort to determine the dominant current directions during deposition of the formation. Pebble and quartz grain orientation as well as heavy mineral and rock composition analyses were made In the laboratory to supplement the field work. -

Cambrian Ordovician
Open File Report LXXVI the shale is also variously colored. Glauconite is generally abundant in the formation. The Eau Claire A Summary of the Stratigraphy of the increases in thickness southward in the Southern Peninsula of Michigan where it becomes much more Southern Peninsula of Michigan * dolomitic. by: The Dresbach sandstone is a fine to medium grained E. J. Baltrusaites, C. K. Clark, G. V. Cohee, R. P. Grant sandstone with well rounded and angular quartz grains. W. A. Kelly, K. K. Landes, G. D. Lindberg and R. B. Thin beds of argillaceous dolomite may occur locally in Newcombe of the Michigan Geological Society * the sandstone. It is about 100 feet thick in the Southern Peninsula of Michigan but is absent in Northern Indiana. The Franconia sandstone is a fine to medium grained Cambrian glauconitic and dolomitic sandstone. It is from 10 to 20 Cambrian rocks in the Southern Peninsula of Michigan feet thick where present in the Southern Peninsula. consist of sandstone, dolomite, and some shale. These * See last page rocks, Lake Superior sandstone, which are of Upper Cambrian age overlie pre-Cambrian rocks and are The Trempealeau is predominantly a buff to light brown divided into the Jacobsville sandstone overlain by the dolomite with a minor amount of sandy, glauconitic Munising. The Munising sandstone at the north is dolomite and dolomitic shale in the basal part. Zones of divided southward into the following formations in sandy dolomite are in the Trempealeau in addition to the ascending order: Mount Simon, Eau Claire, Dresbach basal part. A small amount of chert may be found in and Franconia sandstones overlain by the Trampealeau various places in the formation. -
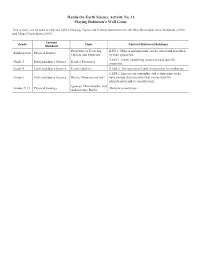
Playing Robinson's Wall Game
Hands On Earth Science Activity No. 13 Playing Robinson’s Wall Game This activity can be used to help teach the following Topics and Content Statements for the Ohio Revised Science Standards (2018) and Model Curriculum (2019): Content Grade Topic Content Statement/Subtopic Standard Properties of Everyday K.PS.1: Objects and materials can be sorted and described Kindergarten Physical Science Objects and Materials by their properties. 3.ESS.1: Earth’s nonliving resources have specific Grade 3 Earth and Space Science Earth’s Resources properties. Grade 4 Earth and Space Science Earth’s Surface 4.ESS.2: The surface of Earth changes due to weathering. 6.ESS.2: Igneous, metamorphic and sedimentary rocks Grade 6 Earth and Space Science Rocks, Minerals and Soil have unique characteristics that can be used for identification and/or classification. Igneous, Metamorphic and Grades 9–12 Physical Geology Multiple connections Sedimentary Rocks No. 13 PLAYING ROBINSON’S WALL GAME by Joseph T. Hannibal, The Cleveland Museum of Natural History There is probably a stone wall somewhere near you. The wall may be a fence or a foundation or other part of a building. Such stone walls can be fascinating geologically, especially if they are constructed of more than one kind of stone. For students at any level, they make excellent places for hands-on dis- covery. Stone fences generally are made with local bedrock or, in glaciated areas, with local glacial erratics. In the case of buildings, many older structures are made of local stone; more recent buildings may be made of more exotic stones imported to the area for a particular project. -

Summary of Hydrogelogic Conditions by County for the State of Michigan. Apple, B.A., and H.W. Reeves 2007. U.S. Geological Surve
In cooperation with the State of Michigan, Department of Environmental Quality Summary of Hydrogeologic Conditions by County for the State of Michigan Open-File Report 2007-1236 U.S. Department of the Interior U.S. Geological Survey Summary of Hydrogeologic Conditions by County for the State of Michigan By Beth A. Apple and Howard W. Reeves In cooperation with the State of Michigan, Department of Environmental Quality Open-File Report 2007-1236 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior DIRK KEMPTHORNE, Secretary U.S. Geological Survey Mark D. Myers, Director U.S. Geological Survey, Reston, Virginia: 2007 For more information about the USGS and its products: Telephone: 1-888-ASK-USGS World Wide Web: http://www.usgs.gov/ Any use of trade, product, or firm names in this publication is for descriptive purposes only and does not imply endorsement by the U.S. Government. Although this report is in the public domain, permission must be secured from the individual copyright owners to reproduce any copyrighted materials contained within this report. Suggested citation Beth, A. Apple and Howard W. Reeves, 2007, Summary of Hydrogeologic Conditions by County for the State of Michi- gan. U.S. Geological Survey Open-File Report 2007-1236, 78 p. Cover photographs Clockwise from upper left: Photograph of Pretty Lake by Gary Huffman. Photograph of a river in winter by Dan Wydra. Photographs of Lake Michigan and the Looking Glass River by Sharon Baltusis. iii Contents Abstract ...........................................................................................................................................................1 -

Late Jurassic Dinosaurs on the Move, Gastroliths and Long-Distance Migration" (2019)
Augustana College Augustana Digital Commons Geography: Student Scholarship & Creative Works Geography Winter 12-8-2019 Late Jurassic Dinosaurs on the Move, Gastroliths and Long- Distance Migration Josh Malone Augustana College, Rock Island Illinois Follow this and additional works at: https://digitalcommons.augustana.edu/geogstudent Part of the Geology Commons, Physical and Environmental Geography Commons, Sedimentology Commons, and the Spatial Science Commons Augustana Digital Commons Citation Malone, Josh. "Late Jurassic Dinosaurs on the Move, Gastroliths and Long-Distance Migration" (2019). Geography: Student Scholarship & Creative Works. https://digitalcommons.augustana.edu/geogstudent/8 This Student Paper is brought to you for free and open access by the Geography at Augustana Digital Commons. It has been accepted for inclusion in Geography: Student Scholarship & Creative Works by an authorized administrator of Augustana Digital Commons. For more information, please contact [email protected]. LATE JURASSIC DINOSAURS ON THE MOVE, GASTROLITHS AND LONG- DISTANCE MIGRATION a senior thesis written by Joshua Malone in partial fulfillment of the graduation requirements for the major in Geography Augustana College Rock Island, Illinois 61201 1 Table of Contents 1. Abstract ................................................................................................................................................ 4 2. Introduction ........................................................................................................................................ -

G-012011-1E Geological Precambrian Timeline Midwest
Copper Harbor Conglomerate Gunflint Formation: Breccia with white quartz Precambrian Geologic Events in the Mid-Continent of North America G-012011-1E 1 inch (Century Mine, Upper Peninsula MI) (Sibley Peninsula, Thunder Bay, ON) Compiled by: Steven D.J. Baumann, Alexandra B. Cory, Micaela M. Krol, Elisa J. Piispa Updated March 2013 Oldest known rock showing a dipole magnetic field: red dacite in Austrailia Paleomagnetic Line 3,800 3,700 3,600 3,500 3,400 3,300 3,200 3,100 3,000 2,900 2,800 2,700 2,600 2,500 2,400 2,300 2,200 2,100 2,000 1,900 1,800 1,700 1,600 1,500 1,400 1,300 1,200 1,100 1,000 900 800 700 600 500 Paleozoic Period Siderian Rhyacian Orosirian Statherian Calymmian Ectasian Stenian Tonian Cryogenian Ediacaran Eoarchean Paleoarchean Mesoarchean Neoarchean Era Paleoproterozoic Mesoproterozoic Neoproterozoic Eon Archean Proterozoic Pass Lake Kama Hill Sibley Group Sediments (Sibley Basin, Thunder Bay Area, ON) McGrath Gneiss McGrath Complex (EC MN) Metamorphic and cataclastic event Formation Formation Outan Island Formation Nipigon Formation Recent Era of Great Mid-continent Basin Formation (MI, IL, IA, IN, KY, MO) 2 inches Marshfield Archean Gneiss (C WI) Linwood Archean Migmatite (C WI) Sudbury Dike Swarm (SE ON) Quinnesec Formation Intrusions (NE WI) Quinnesec Formation Metamorphism (NE WI) Hatfield Gneiss (WC WI) Pre-Quinnesec Formations deposited (NE WI) Upper Rove Formation Baraboo Quartzite LEGEND (Sibley Peninsula, Thunder Bay, ON) Gray granodioritic phase Montevideo Gneiss (SW MN) Red granite phase Montevideo Gneiss -

Paleozoic Stratigraphic Nomenclature for Wisconsin (Wisconsin
UNIVERSITY EXTENSION The University of Wisconsin Geological and Natural History Survey Information Circular Number 8 Paleozoic Stratigraphic Nomenclature For Wisconsin By Meredith E. Ostrom"'" INTRODUCTION The Paleozoic stratigraphic nomenclature shown in the Oronto a Precambrian age and selected the basal contact column is a part of a broad program of the Wisconsin at the top of the uppermost volcanic bed. It is now known Geological and Natural History Survey to re-examine the that the Oronto is unconformable with older rocks in some Paleozoic rocks of Wisconsin and is a response to the needs areas as for example at Fond du Lac, Minnesota, where of geologists, hydrologists and the mineral industry. The the Outer Conglomerate and Nonesuch Shale are missing column was preceded by studies of pre-Cincinnatian cyclical and the younger Freda Sandstone rests on the Thompson sedimentation in the upper Mississippi valley area (Ostrom, Slate (Raasch, 1950; Goldich et ai, 1961). An unconformity 1964), Cambro-Ordovician stratigraphy of southwestern at the upper contact in the Upper Peninsula of Michigan Wisconsin (Ostrom, 1965) and Cambrian stratigraphy in has been postulated by Hamblin (1961) and in northwestern western Wisconsin (Ostrom, 1966). Wisconsin wlle're Atwater and Clement (1935) describe un A major problem of correlation is the tracing of outcrop conformities between flat-lying quartz sandstone (either formations into the subsurface. Outcrop definitions of Mt. Simon, Bayfield, or Hinckley) and older westward formations based chiefly on paleontology can rarely, if dipping Keweenawan volcanics and arkosic sandstone. ever, be extended into the subsurface of Wisconsin because From the above data it would appear that arkosic fossils are usually scarce or absent and their fragments cari rocks of the Oronto Group are unconformable with both seldom be recognized in drill cuttings. -

The End of Midcontinent Rift Magmatism and the Paleogeography of Laurentia
THEMED ISSUE: Tectonics at the Micro- to Macroscale: Contributions in Honor of the University of Michigan Structure-Tectonics Research Group of Ben van der Pluijm and Rob Van der Voo The end of Midcontinent Rift magmatism and the paleogeography of Laurentia Luke M. Fairchild1, Nicholas L. Swanson-Hysell1, Jahandar Ramezani2, Courtney J. Sprain1, and Samuel A. Bowring2 1DEPARTMENT OF EARTH AND PLANETARY SCIENCE, UNIVERSITY OF CALIFORNIA, BERKELEY, CALIFORNIA 94720, USA 2DEPARTMENT OF EARTH, ATMOSPHERIC AND PLANETARY SCIENCES, MASSACHUSETTS INSTITUTE OF TECHNOLOGY, CAMBRIDGE, MASSACHUSETTS 02139, USA ABSTRACT Paleomagnetism of the North American Midcontinent Rift provides a robust paleogeographic record of Laurentia (cratonic North America) from ca. 1110 to 1070 Ma, revealing rapid equatorward motion of the continent throughout rift magmatism. Existing age and paleomagnetic constraints on the youngest rift volcanic and sedimentary rocks have been interpreted to record a slowdown of this motion as rifting waned. We present new paleomagnetic and geochronologic data from the ca. 1090–1083 Ma “late-stage” rift volcanic rocks exposed as the Lake Shore Traps (Michigan), the Schroeder-Lutsen basalts (Minnesota), and the Michipicoten Island Formation (Ontario). The paleomagnetic data allow for the development of paleomagnetic poles for the Schroeder-Lutsen basalts (187.8°E, 27.1°N; A95 = 3.0°, N = 50) and the Michipicoten Island Formation (174.7°E, 17.0°N; A95 = 4.4°, N = 23). Temporal constraints on late-stage paleomagnetic poles are provided by high-precision, 206Pb-238U zircon dates from a Lake Shore Traps andesite (1085.57 ± 0.25 Ma; 2σ internal errors), a Michipicoten Island Formation tuff (1084.35 ± 0.20 Ma) and rhyolite (1083.52 ± 0.23 Ma), and a Silver Bay aplitic dike from the Beaver Bay Complex (1091.61 ± 0.14 Ma), which is overlain by the Schroeder-Lutsen basalt flows. -

Geology of Michigan and the Great Lakes
35133_Geo_Michigan_Cover.qxd 11/13/07 10:26 AM Page 1 “The Geology of Michigan and the Great Lakes” is written to augment any introductory earth science, environmental geology, geologic, or geographic course offering, and is designed to introduce students in Michigan and the Great Lakes to important regional geologic concepts and events. Although Michigan’s geologic past spans the Precambrian through the Holocene, much of the rock record, Pennsylvanian through Pliocene, is miss- ing. Glacial events during the Pleistocene removed these rocks. However, these same glacial events left behind a rich legacy of surficial deposits, various landscape features, lakes, and rivers. Michigan is one of the most scenic states in the nation, providing numerous recre- ational opportunities to inhabitants and visitors alike. Geology of the region has also played an important, and often controlling, role in the pattern of settlement and ongoing economic development of the state. Vital resources such as iron ore, copper, gypsum, salt, oil, and gas have greatly contributed to Michigan’s growth and industrial might. Ample supplies of high-quality water support a vibrant population and strong industrial base throughout the Great Lakes region. These water supplies are now becoming increasingly important in light of modern economic growth and population demands. This text introduces the student to the geology of Michigan and the Great Lakes region. It begins with the Precambrian basement terrains as they relate to plate tectonic events. It describes Paleozoic clastic and carbonate rocks, restricted basin salts, and Niagaran pinnacle reefs. Quaternary glacial events and the development of today’s modern landscapes are also discussed. -

ECONOMIC GEOLOGY of the SAND and SANDSTONE RESOURCES of MICHIGAN
ECONOMIC GEOLOGY of the SAND and SANDSTONE RESOURCES of MICHIGAN Geological Survey Division Report of Investigation 21 By E. William Heinrich Copyright © 2001 by the Michigan Department of Environmental Quality (DEQ) Geological Survey Division (GSD). The DEQ GSD grants permission to publish or reproduce this document, all or in part, for non-profit purposes. The contents of this electronic document (whole or in part) can be used if, and only if, additional fees are not associated with the use or distribution of this document and credit is given to the DEQ GSD and the author(s). This copyright statement must appear in any and all electronic or print documents using this file or any part thereof. Contents of this Report PREFACE ............................................................................4 MARSHALL SANDSTONE ............................................... 20 ABSTRACT..........................................................................4 Grindstone Industry................................................... 20 INTRODUCTION..................................................................4 Napoleon Sandstone................................................. 20 MIDDLE PRECAMBRIAN QUARTZITES ...........................5 Previous Work................................................... 20 General Geology ............................................... 21 General........................................................................5 Quarries............................................................. 21 Mesnard Quartzite .......................................................5 -

Bedrock Maps
1987 BEDROCK GEOLOGY OF MICHIGAN BEDROCK GEOLOGY OF EASTERN UPPER PENINSULA MACKINAC BRECCIA BOIS BLANC FORMATION GARDEN ISLAND FORMATION BASS ISLAND GROUP SALINA GROUP SAINT IGNACE DOLOMITE POINT AUX CHENES SHALE ENGADINE GROUP MANISTIQUE GROUP BURNT BLUFF GROUP KEWEENAW CABOT HEAD SHALE MANITOULIN DOLOMITE QUEENSTON SHALE BIG HILL DOLOMITE HOUGHTON STONINGTON FORMATION UTICA SHALE MEMBER COLLINGWOOD SHALE MEMBER TRENTON GROUP BLACK RIVER GROUP ONTONAGON BARAGA PRAIRIE DU CHIEN GROUP TREMPEALEAU FORMATION MUNISING FORMATION GOGEBIC LU CE MARQUETTE ALGER CHIPPEWA IRON MACKINAC SCHOOLC RAF T DELT A DICKIN SON BEDROCK GEOLOGY OF WESTERN UPPER PENINSULA MACKINAC BRECCIA JACOBSVILLE SANDSTONE EMMET MENOMINEE FREDA SANDSTONE CHEBOYGAN NONESUCH FORMATION PRESQUE ISLE COPPER HARBOR CONGLOMERATE OAK BLUFF FORMATION CHAR LEVOIX PORTAGE LAKE VOLCANICS MONTMORENCY SIEMENS CREEK FORMATION ANT RIM ALPENA INTRUSIVE OTSEGO QUINNESEC FORMATION LEELANAU PAINT RIVER GROUP RIVERTON IRON FORMATION BIJIKI IRON FORMATION GRAND TR AVERSE ALCONA KALKASKA CRAW FOR D OSCOD A NEGAUNEE IRON FORMATION BENZIE IRONWOOD IRON FORMATION DUNN CREEK FORMATION BADWATER GREENSTONE MICHIGAMME FORMATION MANISTEE WEXFORD MISSAUKEE ROSCOMMON OGEMAW IOSCO GOODRICH QUARTZITE HEMLOCK FORMATION BEDROCK GEOLOGY OF ARENAC MENOMINEE & CHOCOLAY GROUPS LOWER PENINSULA EMPEROR VULCANIC COMPLEX MASON LAKE OSCEOLA CLARE GLADWIN SIAMO SLATE & AJIBIK QUARTZITE RED BEDS HURON PALMS FORMATION GRAND RIVER FORMATION CHOCOLAY GROUP SAGINAW FORMATION BAY RANDVILLE DOLOMITE BAYPORT LIMESTONE MICHIGAN -

Contents Illustrations Table Introduction
The Base of the Upper Keweenawan, ILLUSTRATIONS Michigan and Wisconsin FIGURE 1. Distribution of the Copper Harbor Conglomerate and included mafic lava members...........................................2 By WALTER S. WHITE 2. Stratigraphic section of the Copper Harbor Conglomerate..2 CONTRIBUTIONS TO STRATIGRAPHY 3. Longitudinal stratigraphic section of Keweenawan rocks below the base of the Nonesuch Shale............................3 4. Mean directions of magnetization for some Keweenawan GEOLOGICAL SURVEY BULLETIN 1354-F rocks.. ..............................................................................6 A proposal to adopt the top of the Copper Harbor 5. Proposed nomenclature for middle and upper Keweenawan Conglomerate as the base of the upper Keweenawan in rocks ................................................................................9 the Lake Superior region TABLE TABLE 1. Compressional wave velocities (km/sec) for selected Keweenawan stratigraphic units, Lake Superior region ...7 ABSTRACT UNITED STATES GOVERNMENT PRINTING OFFICE, The top of the Copper Harbor Conglomerate (base of WASHINGTON : 1972 Nonesuch Shale) is a more satisfactory boundary between upper and middle Keweenawan rocks in northern Michigan and UNITED STATES DEPARTMENT OF THE INTERIOR adjacent parts of Wisconsin than the various horizons that ROGERS G. B. MORTON, Secretary have been used hitherto without stratigraphic consistency from GEOLOGICAL SURVEY place to place. Irving’s original boundary (1883) cannot be V. E. McKelvey, Director followed away from the Keweenaw Peninsula. The top of the Copper Harbor Conglomerate comes closer to marking the Library of Congress catalog-card No. 72-75175 close of Keweenawan volcanism than other major boundaries and actually adheres more closely to Irving’s original concept For sale by the Superintendent of Documents, U.S. Government Printing Office than the boundary that he himself chose.