Original Article Identification of Differentially Expressed Genes Between Male and Female Patients with Acute Myocardial Infarction Based on Microarray Data
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Small Cell Ovarian Carcinoma: Genomic Stability and Responsiveness to Therapeutics
Gamwell et al. Orphanet Journal of Rare Diseases 2013, 8:33 http://www.ojrd.com/content/8/1/33 RESEARCH Open Access Small cell ovarian carcinoma: genomic stability and responsiveness to therapeutics Lisa F Gamwell1,2, Karen Gambaro3, Maria Merziotis2, Colleen Crane2, Suzanna L Arcand4, Valerie Bourada1,2, Christopher Davis2, Jeremy A Squire6, David G Huntsman7,8, Patricia N Tonin3,4,5 and Barbara C Vanderhyden1,2* Abstract Background: The biology of small cell ovarian carcinoma of the hypercalcemic type (SCCOHT), which is a rare and aggressive form of ovarian cancer, is poorly understood. Tumourigenicity, in vitro growth characteristics, genetic and genomic anomalies, and sensitivity to standard and novel chemotherapeutic treatments were investigated in the unique SCCOHT cell line, BIN-67, to provide further insight in the biology of this rare type of ovarian cancer. Method: The tumourigenic potential of BIN-67 cells was determined and the tumours formed in a xenograft model was compared to human SCCOHT. DNA sequencing, spectral karyotyping and high density SNP array analysis was performed. The sensitivity of the BIN-67 cells to standard chemotherapeutic agents and to vesicular stomatitis virus (VSV) and the JX-594 vaccinia virus was tested. Results: BIN-67 cells were capable of forming spheroids in hanging drop cultures. When xenografted into immunodeficient mice, BIN-67 cells developed into tumours that reflected the hypercalcemia and histology of human SCCOHT, notably intense expression of WT-1 and vimentin, and lack of expression of inhibin. Somatic mutations in TP53 and the most common activating mutations in KRAS and BRAF were not found in BIN-67 cells by DNA sequencing. -

Genomic Correlates of Relationship QTL Involved in Fore- Versus Hind Limb Divergence in Mice
Loyola University Chicago Loyola eCommons Biology: Faculty Publications and Other Works Faculty Publications 2013 Genomic Correlates of Relationship QTL Involved in Fore- Versus Hind Limb Divergence in Mice Mihaela Palicev Gunter P. Wagner James P. Noonan Benedikt Hallgrimsson James M. Cheverud Loyola University Chicago, [email protected] Follow this and additional works at: https://ecommons.luc.edu/biology_facpubs Part of the Biology Commons Recommended Citation Palicev, M, GP Wagner, JP Noonan, B Hallgrimsson, and JM Cheverud. "Genomic Correlates of Relationship QTL Involved in Fore- Versus Hind Limb Divergence in Mice." Genome Biology and Evolution 5(10), 2013. This Article is brought to you for free and open access by the Faculty Publications at Loyola eCommons. It has been accepted for inclusion in Biology: Faculty Publications and Other Works by an authorized administrator of Loyola eCommons. For more information, please contact [email protected]. This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 License. © Palicev et al., 2013. GBE Genomic Correlates of Relationship QTL Involved in Fore- versus Hind Limb Divergence in Mice Mihaela Pavlicev1,2,*, Gu¨ nter P. Wagner3, James P. Noonan4, Benedikt Hallgrı´msson5,and James M. Cheverud6 1Konrad Lorenz Institute for Evolution and Cognition Research, Altenberg, Austria 2Department of Pediatrics, Cincinnati Children‘s Hospital Medical Center, Cincinnati, Ohio 3Yale Systems Biology Institute and Department of Ecology and Evolutionary Biology, Yale University 4Department of Genetics, Yale University School of Medicine 5Department of Cell Biology and Anatomy, The McCaig Institute for Bone and Joint Health and the Alberta Children’s Hospital Research Institute for Child and Maternal Health, University of Calgary, Calgary, Canada 6Department of Anatomy and Neurobiology, Washington University *Corresponding author: E-mail: [email protected]. -
Primepcr™Assay Validation Report
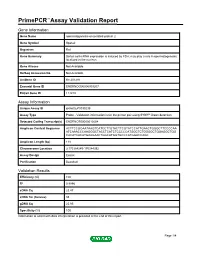
PrimePCR™Assay Validation Report Gene Information Gene Name spermatogenesis-associated protein 2 Gene Symbol Spata2 Organism Rat Gene Summary Sertoli cell mRNA expression is induced by FSH; may play a role in spermatogenesis; localized to the nucleus Gene Aliases Not Available RefSeq Accession No. Not Available UniGene ID Rn.201291 Ensembl Gene ID ENSRNOG00000009207 Entrez Gene ID 114210 Assay Information Unique Assay ID qRnoCEP0030238 Assay Type Probe - Validation information is for the primer pair using SYBR® Green detection Detected Coding Transcript(s) ENSRNOT00000012604 Amplicon Context Sequence ACTTCCGGAATAAGTCATCCTTGTACTTCGTATCCATTGAACTGGGCTTCCCCAA ATCAAACCCAAGGGCTACCTCATCTCCCCCATGGCTCTGGGGCTGGAGGCTGG CACATCACATGAAGAACTGGCATGGTGCCCATGGACCAGC Amplicon Length (bp) 118 Chromosome Location 3:170384245-170384392 Assay Design Exonic Purification Desalted Validation Results Efficiency (%) 100 R2 0.9996 cDNA Cq 23.47 cDNA Tm (Celsius) 85 gDNA Cq 25.95 Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 1/4 PrimePCR™Assay Validation Report Spata2, Rat Amplification Plot Amplification of cDNA generated from 25 ng of universal reference RNA Melt Peak Melt curve analysis of above amplification Standard Curve Standard curve generated using 20 million copies of template diluted 10-fold to 20 copies Page 2/4 PrimePCR™Assay Validation Report Products used to generate validation data Real-Time PCR Instrument CFX384 Real-Time PCR Detection System Reverse Transcription Reagent iScript™ Advanced cDNA Synthesis Kit for RT-qPCR Real-Time PCR Supermix SsoAdvanced™ SYBR® Green Supermix Experimental Sample qPCR Reference Total RNA Data Interpretation Unique Assay ID This is a unique identifier that can be used to identify the assay in the literature and online. Detected Coding Transcript(s) This is a list of the Ensembl transcript ID(s) that this assay will detect. -

Recombinant Rabbit Anti-Human RAB8A Monoclonal Antibody, Clone NKG-S33-80-4 (CABT- Z616R) This Product Is for Research Use Only and Is Not Intended for Diagnostic Use
Recombinant Rabbit Anti-Human RAB8A Monoclonal Antibody, clone NKG-S33-80-4 (CABT- Z616R) This product is for research use only and is not intended for diagnostic use. PRODUCT INFORMATION Immunogen Recombinant full length protein within Human RAB8A aa 1 to the C-terminus. Isotype IgG Source/Host Rabbit Species Reactivity Human Clone NKG-S33-80-4 Purification Protein A purified Conjugate Unconjugated Applications WB, IHC-P, ICC/IF, FC, IP Recommended dilution: WB: 1:1000 IHC-P: 1:1000 ICC/IF: 1:500 FC: 1:600 IP: 1:30 Positive Control WB: HEK-293T, HCT116 and HeLa whole cell lysates. IHC-P: Human bladder cancer and breast tissues. ICC/IF: A549 and HeLa cells. Flow Cyt: A549 cells. IP: A549 whole cell lysate. Format Liquid Concentration Lot specific Size 100 μl Buffer PBS, 40% Glycerol (glycerin, glycerine), 0.05% BSA, pH 7.2. Preservative 0.01% Sodium azide Storage Store at +4℃ short term (1-2 weeks). Upon delivery aliquot. Store at -20℃. Stable for 12 months at -20℃. 45-1 Ramsey Road, Shirley, NY 11967, USA Email: [email protected] Tel: 1-631-624-4882 Fax: 1-631-938-8221 1 © Creative Diagnostics All Rights Reserved Ship Wet ice Warnings This product is for research use only and is not intended for diagnostic use. BACKGROUND Introduction RAB8A may be involved in vesicular trafficking and neurotransmitter release. Together with RAB11A, RAB3IP, the exocyst complex, PARD3, PRKCI, ANXA2, CDC42 and DNMBP promotes transcytosis of PODXL to the apical membrane initiation sites (AMIS), apical surface formation and lumenogenesis. Together with MYO5B and RAB11A participates in epithelial cell polarization. -

The Autoimmune Signature of Hyperinflammatory Multisystem Inflammatory Syndrome in Children Rebecca A. Porritt1,*, Aleksandra Bi
The autoimmune signature of hyperinflammatory multisystem inflammatory syndrome in children Rebecca A. Porritt1,*, Aleksandra Binek2,*, Lisa Paschold3,*, Magali Noval Rivas1, Angela Mc Ardle2, Lael M. Yonker4,5, Galit Alter4,5,6, Harsha Chandnani7, Merrick Lopez7, Alessio Fasano4,5, Jennifer E. Van Eyk2,8,†, Mascha Binder3,† and Moshe Arditi1,9,† 1Departments of Pediatrics, Division of Infectious Diseases and Immunology, and Infectious and Immunologic Diseases Research Center (IIDRC), Department of Biomedical Sciences, Cedars- Sinai Medical Center, Los Angeles, CA, USA. 2Advanced Clinical Biosystems Research Institute, The Smidt Heart Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. 3Department of Internal Medicine IV, Oncology/Hematology, Martin-Luther-University Halle- Wittenberg, 06120 Halle (Saale), Germany 4Massachusetts General Hospital, Mucosal Immunology and Biology Research Center and Department of Pediatrics, Boston, MA, USA. 5Harvard Medical School, Boston, MA, USA. 6Ragon Institute of MIT, MGH and Harvard, Cambridge, MA, USA. 7Department of Pediatrics, Loma Linda University Hospital, CA, USA. 8Barbra Streisand Women's Heart Center, Cedars-Sinai Smidt Heart Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA. 9Cedars-Sinai Smidt Heart Institute, Cedars-Sinai Medical Center, Los Angeles, CA, USA * these authors contributed equally † these senior authors contributed equally Corresponding author: Moshe Arditi, MD 8700 Beverly Blvd. Davis Building, Rooms D4024, 4025, 4027 Los Angeles, CA 90048 Phone:310-423-4471 -

A Computational Approach for Defining a Signature of Β-Cell Golgi Stress in Diabetes Mellitus
Page 1 of 781 Diabetes A Computational Approach for Defining a Signature of β-Cell Golgi Stress in Diabetes Mellitus Robert N. Bone1,6,7, Olufunmilola Oyebamiji2, Sayali Talware2, Sharmila Selvaraj2, Preethi Krishnan3,6, Farooq Syed1,6,7, Huanmei Wu2, Carmella Evans-Molina 1,3,4,5,6,7,8* Departments of 1Pediatrics, 3Medicine, 4Anatomy, Cell Biology & Physiology, 5Biochemistry & Molecular Biology, the 6Center for Diabetes & Metabolic Diseases, and the 7Herman B. Wells Center for Pediatric Research, Indiana University School of Medicine, Indianapolis, IN 46202; 2Department of BioHealth Informatics, Indiana University-Purdue University Indianapolis, Indianapolis, IN, 46202; 8Roudebush VA Medical Center, Indianapolis, IN 46202. *Corresponding Author(s): Carmella Evans-Molina, MD, PhD ([email protected]) Indiana University School of Medicine, 635 Barnhill Drive, MS 2031A, Indianapolis, IN 46202, Telephone: (317) 274-4145, Fax (317) 274-4107 Running Title: Golgi Stress Response in Diabetes Word Count: 4358 Number of Figures: 6 Keywords: Golgi apparatus stress, Islets, β cell, Type 1 diabetes, Type 2 diabetes 1 Diabetes Publish Ahead of Print, published online August 20, 2020 Diabetes Page 2 of 781 ABSTRACT The Golgi apparatus (GA) is an important site of insulin processing and granule maturation, but whether GA organelle dysfunction and GA stress are present in the diabetic β-cell has not been tested. We utilized an informatics-based approach to develop a transcriptional signature of β-cell GA stress using existing RNA sequencing and microarray datasets generated using human islets from donors with diabetes and islets where type 1(T1D) and type 2 diabetes (T2D) had been modeled ex vivo. To narrow our results to GA-specific genes, we applied a filter set of 1,030 genes accepted as GA associated. -

The LRRC8A:C Heteromeric Channel Is a Cgamp Transporter and the Dominant
bioRxiv preprint doi: https://doi.org/10.1101/2020.02.13.948273; this version posted February 14, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. The LRRC8A:C Heteromeric Channel Is a cGAMP Transporter and the Dominant cGAMP Importer in Human Vasculature Cells Lauren J. Lahey1,2, Xianlan Wen3, Rachel E. Mardjuki2,4, Volker Böhnert2,5, Christopher Ritchie2,5, Jacqueline A. Carozza2,4, Gaelen T. Hess2,6, Merritt Maduke3, Michael C. Bassik2,6, and Lingyin Li2,5,7,* 1Biophysics Program, 2Stanford ChEM-H, 3Department of Molecular and Cellular Physiology, 4Department of Chemistry, 5Department of Biochemistry, 6Department of Genetics, Stanford University School of Medicine, Stanford, CA 94305 USA. 7Lead Contact *Correspondence: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/2020.02.13.948273; this version posted February 14, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. SUMMARY Extracellular 2'3'-cyclic-GMP-AMP (cGAMP) is an immunotransmitter secreted by cancer cells and taken up by host cells to activate the anti-cancer STING pathway. No cGAMP exporter has been identified, and SLC19A1, a recently identified cGAMP importer, does not account for the import actiVity in most cell types. Here, we identify the LRRC8A:C heteromeric channel, a volume-regulated anion channel (VRAC), as a cGAMP transporter. This channel mediates cGAMP import or export depending on the cGAMP chemical gradient. cGAMP influences anion influx through VRAC, indicating it is likely a direct substrate of the channel. -

Bppart and Bpmax: RNA-RNA Interaction Partition Function and Structure Prediction for the Base Pair Counting Model
BPPart and BPMax: RNA-RNA Interaction Partition Function and Structure Prediction for the Base Pair Counting Model Ali Ebrahimpour-Boroojeny, Sanjay Rajopadhye, and Hamidreza Chitsaz ∗ Department of Computer Science, Colorado State University Abstract A few elite classes of RNA-RNA interaction (RRI), with complex roles in cellular functions such as miRNA-target and lncRNAs in human health, have already been studied. Accordingly, RRI bioinfor- matics tools tailored for those elite classes have been proposed in the last decade. Interestingly, there are somewhat unnoticed mRNA-mRNA interactions in the literature with potentially drastic biological roles. Hence, there is a need for high-throughput generic RRI bioinformatics tools. We revisit our RRI partition function algorithm, piRNA, which happens to be the most comprehensive and computationally-intensive thermodynamic model for RRI. We propose simpler models that are shown to retain the vast majority of the thermodynamic information that piRNA captures. We simplify the energy model and instead consider only weighted base pair counting to obtain BPPart for Base-pair Partition function and BPMax for Base-pair Maximization which are 225 and 1350 faster ◦ × × than piRNA, with a correlation of 0.855 and 0.836 with piRNA at 37 C on 50,500 experimentally charac- terized RRIs. This correlation increases to 0.920 and 0.904, respectively, at 180◦C. − Finally, we apply our algorithm BPPart to discover two disease-related RNAs, SNORD3D and TRAF3, and hypothesize their potential roles in Parkinson's disease and Cerebral Autosomal Dominant Arteri- opathy with Subcortical Infarcts and Leukoencephalopathy (CADASIL). 1 Introduction Since mid 1990s with the advent of RNA interference discovery, RNA-RNA interaction (RRI) has moved to the spotlight in modern, post-genome biology. -

TTDN1 (MPLKIP) (NM 138701) Human 3' UTR Clone Product Data
OriGene Technologies, Inc. 9620 Medical Center Drive, Ste 200 Rockville, MD 20850, US Phone: +1-888-267-4436 [email protected] EU: [email protected] CN: [email protected] Product datasheet for SC204348 TTDN1 (MPLKIP) (NM_138701) Human 3' UTR Clone Product data: Product Type: 3' UTR Clones Product Name: TTDN1 (MPLKIP) (NM_138701) Human 3' UTR Clone Vector: pMirTarget (PS100062) Symbol: MPLKIP Synonyms: ABHS; C7orf11; ORF20; TTD4 ACCN: NM_138701 Insert Size: 2000 bp This product is to be used for laboratory only. Not for diagnostic or therapeutic use. View online » ©2021 OriGene Technologies, Inc., 9620 Medical Center Drive, Ste 200, Rockville, MD 20850, US 1 / 3 TTDN1 (MPLKIP) (NM_138701) Human 3' UTR Clone – SC204348 Insert Sequence: >SC204348 3’UTR clone of NM_138701 The sequence shown below is from the reference sequence of NM_138701. The complete sequence of this clone may contain minor differences, such as SNPs. Blue=Stop Codon Red=Cloning site GGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGAAAGATCGCCGTG TAACAATTGGCAGAGCTCAGAATTCAAGCGATCGCC ACAGGCAAAAAAGGAAGATACTTTTGTTAACATTTCTGAAATTCAACTGGAAGCTTCATGTGTCAGGAA CATCTTGGACAAAACTTTAAGTTGTGTTGATATAAATTTACCCAAAGATGATGACTTTGATTGGATAAT TAGTAAGGTCTTTTTGTTATTTTTCATCGTATCAGGTATTGTTGATATTAGAGAAAAAAGTAGGATAAC TTGCAACATTTAGCTCTGGAAGTACCTACCACATTTTAGAGATTTACCGTTTCCATATATTTAACATTC CTGGTTACATAATGGACATTTGTCTTTTAATGTTTTTTCAATGTTTTAAAATAAAACATTTTGTCTTCT AGCTATTGTGGTTTTGTGGTATGATAAAGAAGTAGACTTACTACAGTAATGCTTTGTAGTCACTTAGAG TTCATAGGTAAATGTTTTGCAAATTATTTTTGAAAATGAAATAGGTAAACCATCCTTTGAGCTGTAGAC -

Selective Transport of Neurotransmitters and Modulators by Distinct Volume-Regulated LRRC8 Anion Channels Darius Lutter1,2,3, Florian Ullrich1,2, Jennifer C
© 2017. Published by The Company of Biologists Ltd | Journal of Cell Science (2017) 130, 1122-1133 doi:10.1242/jcs.196253 RESEARCH ARTICLE Selective transport of neurotransmitters and modulators by distinct volume-regulated LRRC8 anion channels Darius Lutter1,2,3, Florian Ullrich1,2, Jennifer C. Lueck1,2,3, Stefan Kempa2 and Thomas J. Jentsch1,2,4,* ABSTRACT volume regulation (Jentsch, 2016; Nilius et al., 1997; Pedersen In response to swelling, mammalian cells release chloride and et al., 2016). This plasma membrane channel, which appears to be organic osmolytes through volume-regulated anion channels expressed in all cells, is normally closed but can be activated by cell swelling or other cellular signaling cascades. In most cells, VRAC (VRACs). VRACs are heteromers of LRRC8A and other LRRC8 − isoforms (LRRC8B to LRRC8E), which are co-expressed in HEK293 opening leads to an efflux of Cl , which, together with an efflux of + + and most other cells. The spectrum of VRAC substrates and its K through independent K channels, lowers the intracellular dependence on particular LRRC8 isoforms remains largely unknown. osmolarity and, by secondarily driving water efflux, results in We show that, besides the osmolytes taurine and myo-inositol, regulatory volume decrease. Cell swelling also stimulates an efflux ‘ ’ myo LRRC8 channels transport the neurotransmitters glutamate, of organic osmolytes such as -inositol, taurine and glutamate. aspartate and γ-aminobutyric acid (GABA) and the co-activator It was controversial whether this release occurs through VRAC or D-serine. HEK293 cells engineered to express defined subsets of through a distinct channel named VSOAC (for volume-sensitive LRRC8 isoforms were used to elucidate the subunit-dependence organic osmolyte/anion channel) (Jackson and Strange, 1993; of transport. -

S41467-020-18249-3.Pdf
ARTICLE https://doi.org/10.1038/s41467-020-18249-3 OPEN Pharmacologically reversible zonation-dependent endothelial cell transcriptomic changes with neurodegenerative disease associations in the aged brain Lei Zhao1,2,17, Zhongqi Li 1,2,17, Joaquim S. L. Vong2,3,17, Xinyi Chen1,2, Hei-Ming Lai1,2,4,5,6, Leo Y. C. Yan1,2, Junzhe Huang1,2, Samuel K. H. Sy1,2,7, Xiaoyu Tian 8, Yu Huang 8, Ho Yin Edwin Chan5,9, Hon-Cheong So6,8, ✉ ✉ Wai-Lung Ng 10, Yamei Tang11, Wei-Jye Lin12,13, Vincent C. T. Mok1,5,6,14,15 &HoKo 1,2,4,5,6,8,14,16 1234567890():,; The molecular signatures of cells in the brain have been revealed in unprecedented detail, yet the ageing-associated genome-wide expression changes that may contribute to neurovas- cular dysfunction in neurodegenerative diseases remain elusive. Here, we report zonation- dependent transcriptomic changes in aged mouse brain endothelial cells (ECs), which pro- minently implicate altered immune/cytokine signaling in ECs of all vascular segments, and functional changes impacting the blood–brain barrier (BBB) and glucose/energy metabolism especially in capillary ECs (capECs). An overrepresentation of Alzheimer disease (AD) GWAS genes is evident among the human orthologs of the differentially expressed genes of aged capECs, while comparative analysis revealed a subset of concordantly downregulated, functionally important genes in human AD brains. Treatment with exenatide, a glucagon-like peptide-1 receptor agonist, strongly reverses aged mouse brain EC transcriptomic changes and BBB leakage, with associated attenuation of microglial priming. We thus revealed tran- scriptomic alterations underlying brain EC ageing that are complex yet pharmacologically reversible. -

WO 2019/079361 Al 25 April 2019 (25.04.2019) W 1P O PCT
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization I International Bureau (10) International Publication Number (43) International Publication Date WO 2019/079361 Al 25 April 2019 (25.04.2019) W 1P O PCT (51) International Patent Classification: CA, CH, CL, CN, CO, CR, CU, CZ, DE, DJ, DK, DM, DO, C12Q 1/68 (2018.01) A61P 31/18 (2006.01) DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, C12Q 1/70 (2006.01) HR, HU, ID, IL, IN, IR, IS, JO, JP, KE, KG, KH, KN, KP, KR, KW, KZ, LA, LC, LK, LR, LS, LU, LY, MA, MD, ME, (21) International Application Number: MG, MK, MN, MW, MX, MY, MZ, NA, NG, NI, NO, NZ, PCT/US2018/056167 OM, PA, PE, PG, PH, PL, PT, QA, RO, RS, RU, RW, SA, (22) International Filing Date: SC, SD, SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, 16 October 2018 (16. 10.2018) TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. (25) Filing Language: English (84) Designated States (unless otherwise indicated, for every kind of regional protection available): ARIPO (BW, GH, (26) Publication Language: English GM, KE, LR, LS, MW, MZ, NA, RW, SD, SL, ST, SZ, TZ, (30) Priority Data: UG, ZM, ZW), Eurasian (AM, AZ, BY, KG, KZ, RU, TJ, 62/573,025 16 October 2017 (16. 10.2017) US TM), European (AL, AT, BE, BG, CH, CY, CZ, DE, DK, EE, ES, FI, FR, GB, GR, HR, HU, ΓΕ , IS, IT, LT, LU, LV, (71) Applicant: MASSACHUSETTS INSTITUTE OF MC, MK, MT, NL, NO, PL, PT, RO, RS, SE, SI, SK, SM, TECHNOLOGY [US/US]; 77 Massachusetts Avenue, TR), OAPI (BF, BJ, CF, CG, CI, CM, GA, GN, GQ, GW, Cambridge, Massachusetts 02139 (US).