Publicación Anticipada Early View
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ecuador's Biodiversity Hotspots
Ecuador’s Biodiversity Hotspots Destination: Andes, Amazon & Galapagos Islands, Ecuador Duration: 19 Days Dates: 29th June – 17th July 2018 Exploring various habitats throughout the wonderful & diverse country of Ecuador Spotting a huge male Andean bear & watching as it ripped into & fed on bromeliads Watching a Eastern olingo climbing the cecropia from the decking in Wildsumaco Seeing ~200 species of bird including 33 species of dazzling hummingbirds Watching a Western Galapagos racer hunting, catching & eating a Marine iguana Incredible animals in the Galapagos including nesting flightless cormorants 36 mammal species including Lowland paca, Andean bear & Galapagos fur seals Watching the incredible and tiny Pygmy marmoset in the Amazon near Sacha Lodge Having very close views of 8 different Andean condors including 3 on the ground Having Galapagos sea lions come up & interact with us on the boat and snorkelling Tour Leader / Guides Overview Martin Royle (Royle Safaris Tour Leader) Gustavo (Andean Naturalist Guide) Day 1: Quito / Puembo Francisco (Antisana Reserve Guide) Milton (Cayambe Coca National Park Guide) ‘Campion’ (Wildsumaco Guide) Day 2: Antisana Wilmar (Shanshu), Alex and Erica (Amazonia Guides) Gustavo (Galapagos Islands Guide) Days 3-4: Cayambe Coca Participants Mr. Joe Boyer Days 5-6: Wildsumaco Mrs. Rhoda Boyer-Perkins Day 7: Quito / Puembo Days 8-10: Amazon Day 11: Quito / Puembo Days 12-18: Galapagos Day 19: Quito / Puembo Royle Safaris – 6 Greenhythe Rd, Heald Green, Cheshire, SK8 3NS – 0845 226 8259 – [email protected] Day by Day Breakdown Overview Ecuador may be a small country on a map, but it is one of the richest countries in the world in terms of life and biodiversity. -

Gallina & Mandujano
Mongabay.com Open Access Journal - Tropical Conservation Science Vol. 2 (2):116-127, 2009 Special issue: introduction Research on ecology, conservation and management of wild ungulates in Mexico Sonia Gallina1 and Salvador Mandujano1 1 Departamento de Biodiversidad y Ecología Animal, Instituto de Ecología A. C., km. 2.5 Carret. Ant. Coatepec No. 351, Congregación del Haya, Xalapa 91070, Ver. México. E‐mail: <[email protected]>; <[email protected]> Abstract This special issue of Tropical Conservation Science provides a synopsis of nine of the eleven presentations on ungulates presented at the Symposium on Ecology and Conservation of Ungulates in Mexico during the Mexican Congress of Ecology held in November 2008 in Merida, Yucatan. Of the eleven species of wild ungulates in Mexico (Baird´s tapir Tapirus bairdii, pronghorn antelope Antilocapra americana, American bison Bison bison, bighorn sheep Ovis canadensis, elk Cervus canadensis, red brocket deer Mazama temama, Yucatan brown brocket Mazama pandora, mule deer Odocoileus hemionus, white-tailed deer Odocoileus virginianus, white-lipped peccary Tayassu pecari and collared peccary Pecari tajacu), studies which concern four of these species are presented: Baird’s tapir and the white lipped peccary, which are tropical species in danger of extinction; the bighorn sheep, of high value for hunting in the north-west; and the white-tailed deer, the most studied ungulate in Mexico due to its wide distribution in the country and high hunting and cultural value. In addition, two studies of exotic species, wild boar (Sus scrofa) and red deer (Cervus elaphus), are presented. Issues addressed in these studies are: population estimates, habitat use, evaluation of UMA (Spanish acronym for ‘Wildlife Conservation, Management and Sustainable Utilization Units’) and ANP (Spanish acronym for ‘Natural Protected Areas’) to sustain minimum viable populations, and the effect of alien species in protected areas and UMA, all of which allow an insight into ungulate conservation and management within the country. -

Canada, Decembre 2008 Library and Bibliotheque Et 1*1 Archives Canada Archives Canada Published Heritage Direction Du Branch Patrimoine De I'edition
ORGANISATION SOCIALE, DYNAMIQUE DE POPULATION, ET CONSERVATION DU CERF HUEMUL (HIPPOCAMELUS BISULCUS) DANS LA PATAGONIE DU CHILI par Paulo Corti these presente au Departement de biologie en vue de l'obtention du grade de docteur es sciences (Ph.D.) FACULTE DES SCIENCES UNIVERSITE DE SHERBROOKE Sherbrooke, Quebec, Canada, decembre 2008 Library and Bibliotheque et 1*1 Archives Canada Archives Canada Published Heritage Direction du Branch Patrimoine de I'edition 395 Wellington Street 395, rue Wellington Ottawa ON K1A0N4 Ottawa ON K1A0N4 Canada Canada Your file Votre reference ISBN: 978-0-494-48538-5 Our file Notre reference ISBN: 978-0-494-48538-5 NOTICE: AVIS: The author has granted a non L'auteur a accorde une licence non exclusive exclusive license allowing Library permettant a la Bibliotheque et Archives and Archives Canada to reproduce, Canada de reproduire, publier, archiver, publish, archive, preserve, conserve, sauvegarder, conserver, transmettre au public communicate to the public by par telecommunication ou par Plntemet, prefer, telecommunication or on the Internet, distribuer et vendre des theses partout dans loan, distribute and sell theses le monde, a des fins commerciales ou autres, worldwide, for commercial or non sur support microforme, papier, electronique commercial purposes, in microform, et/ou autres formats. paper, electronic and/or any other formats. The author retains copyright L'auteur conserve la propriete du droit d'auteur ownership and moral rights in et des droits moraux qui protege cette these. this thesis. Neither the thesis Ni la these ni des extraits substantiels de nor substantial extracts from it celle-ci ne doivent etre imprimes ou autrement may be printed or otherwise reproduits sans son autorisation. -
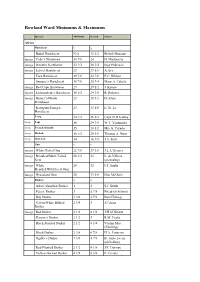
Rowland Ward Minimums & Maximums
Rowland Ward Minimums & Maximums Species Minimum Record Owner Africa Hartebeest x x x Bubal Hartebeest N/A 15 1/2 British Museum image Coke’s Hartebeest 18 7/8 24 D. Mackenzie image Western Hartebeest 22 1/2 28 3/4 Inge Pederson image Lelwel Hartebeest 23 27 5/8 A.Avy Tora Hartebeest 19 7/8 22 7/8 F.C. Hibben Swayne’s Hartebeest 16 7/8 20 3/4 Mary A. Cabela image Red/Cape Hartebeest 23 29 1/2 J. Krause image Lichtenstein’s Hartebeest 18 1/2 24 3/8 R. Rohwer image Hunter’s/Hirola 23 28 1/2 H. Klein Hartebeest Korrigum/Senegal 23 33 1/8 L. St. Lo Hartebeest Tiang 20 1/2 26 1/2 Capt. D.H Gawne image Topi 16 24 3/8 W.T. Yoshimoto image Tsessebe/Sassaby 15 18 1/2 Mrs A. Curado image Blesbok 16 1/2 20 5/8 Thomas A. Hunt image Bontebok 14 16 3/8 J.A. Feist Gnu x x x image White-Tailed Gnu 22 7/8 29 3/8 J.L. L’Ecuyer image Brindled/Black-Tailed 28 1/2 36 L. de Villiers Gnu (picked up) image White 28 32 I.T. Smith Bearded/Wildebeest Gnu image Nyasaland Gnu 28 33 1/8 Don McGuirt Duikers x x x Aders’/Zanzibar Duiker 1 2 S.J. Smith Peters’ Duiker 3 5 7/8 Friedrich Schmitt Bay Duiker 2 3/8 4 7/8 Bela Hidvegi Gabon/White Bellied 2 3/4 5 J.Cousin Duiker image Red Duiker 2 1/2 4 1/8 J.H.M Niblett Harvey’s Duiker 2 1/2 5 R.M. -

Tapirus Pinchaque
ISSN 1813-2286 Volume 22 • No. 31 December 2013 TAPIR CONSERVATION The Newsletter of the IUCN/SSC Tapir Specialist Group www.tapirs.org Printing and distribution of the Tapir Conservation Newsletter is supported by the Houston Zoo Inc., 1513 N. Mac Gregor, Houston, Texas 77030, United States, http://www.houstonzoo.org 2 THE NEWSLETTER OF THE IUCN/SSC TAPIR SPECIALIST GROUP CONTENTS TAPIR CONSERVATION Volume 22 No. 31 Dec 2013 Abbreviation Tapir Cons. From the Chair 3 ISSN 1813-2286 Letter from the Chair Patrícia Medici 3 Website www.tapirs.org Conservation 5 Contributions Anders Gonçalves da Silva (Australia) Editor E-mail: [email protected] Camera-trap Records of Mountain Tapir in Puracé National Park, Colombia Sebastián Duque López, Melissa Abud, Humberto Calero, Layout & Danielle Lalonde (Australia) Stephany Valderrama 5 Distribution Editors Kelly J. Russo (United States) First Report of Positive Serological Response to the Hemoparasite, Babesia caballi, in Mountain Tapir Armando X. Castellanos P. 9 Editorial Board Patrícia Medici E-mail: [email protected]; [email protected] Tapir Conservation Trundles Forward in Belize Carl Traeholt (Denmark/Malaysia) Jamal Andrewin-Bohn 10 Mathias Tobler (Switzerland/Peru) Conservation Medicine 12 Anders Gonçalves da Silva (Australia) Identifying an Effective Treatment for Corneal Ulceration in Diego J. Lizcano (Colombia) Captive Tapirs Mari-Ann O. Da Silva, Catalina Hermoza, Gianmarco Rojas, Matthew Colbert (United States) J. Michelle Freundt 12 Budhan Pukazhenthi (United States) Immobilization of Baird’s Tapir (Tapirus bairdii) Using Benoit de Thoisy (French Guiana) Thiafenthanil Oxalate (A-3080) in Combination with Xylazine and Ketamine Stefan Seitz (Germany) Jonathan Pérez Flores 15 Production This issue is kindly sponsored by Houston Zoo Inc., Contributions 20 & Distribution Kelly Russo, 1513 North Mac Gregor, Houston, Texas 77030, USA. -

Saving the Symbol of the Andes: a Range Wide Conservation Priority Setting Exercise for the Andean Condor (Vultur Gryphus)
Saving the Symbol of the Andes: A Range Wide Conservation Priority Setting Exercise for the Andean Condor (Vultur gryphus) Robert Wallace, Ariel Reinaga, Natalia Piland, Renzo Piana, F. Hernán Vargas, Rosa Elena Zegarra, Pablo Alarcón, Sergio Alvarado, José Álvarez, Fernando Angulo, Vanesa Astore, Francisco Ciri, Jannet Cisneros, Celeste Cóndor, Víctor Escobar, Martín Funes, Jessica Gálvez-Durand, Carolina Gargiulo, Sandra Gordillo, Javier Heredia, Sebastián Kohn, Alejandro Kusch, Sergio Lambertucci, Diego Méndez, Rubén Morales, Alexander More, Adrián Naveda- Rodríguez, David Oehler, Oscar Ospina-Herrera, Andrés Ortega, José Antonio Otero, Fausto Sáenz-Jiménez, Carlos Silva, Claudia Silva, Rosa Vento, Guillermo Wiemeier, Galo Zapata-Ríos & Lorena Zurita May 2020 Results of Andean Condor Range Wide Priority Setting Workshop, May 6th-9th 2015, held in Lima, Peru. Author order is based on the following criteria. The first author led the design of the workshop and research, raised funding for the meeting, provided data and supervised the systematization of Andean condor data across the range, facilitated the RWPS portion of the workshop, and led write-up efforts. The second author systematized Andean condor data and produced maps and spatial analyses for the workshop and the publication. The third author produced a note record of the workshop and then wrote up significant portions of the document. The next three authors were fundamental in the organization and design of the workshop, provided data and reviewed and contributed to this document. The next 31 authors participated in the workshop, generously provided data, and reviewed and edited the report. 1 Saving the Symbol of the Andes: A Range Wide Conservation Priority Setting Exercise for the Andean Condor (Vultur gryphus) Thomas Kramer4 5 Title: Saving the Symbol of the Andes: A Range Wide Conservation Priority Setting Exercise for the Andean Condor (Vultur gryphus) First edition: May 2020 Editor: Robert B. -
Large Mammals of the High Andes a Survey of Community Composition and Habitat Usage in the Highlands of Angochagua Sage Fox SIT Study Abroad
SIT Graduate Institute/SIT Study Abroad SIT Digital Collections Independent Study Project (ISP) Collection SIT Study Abroad Fall 2018 Large Mammals of the High Andes A Survey of Community Composition and Habitat Usage in the Highlands of Angochagua Sage Fox SIT Study Abroad Follow this and additional works at: https://digitalcollections.sit.edu/isp_collection Part of the Animal Studies Commons, Biodiversity Commons, Latin American Studies Commons, and the Zoology Commons Recommended Citation Fox, Sage, "Large Mammals of the High Andes A Survey of Community Composition and Habitat Usage in the Highlands of Angochagua" (2018). Independent Study Project (ISP) Collection. 2925. https://digitalcollections.sit.edu/isp_collection/2925 This Unpublished Paper is brought to you for free and open access by the SIT Study Abroad at SIT Digital Collections. It has been accepted for inclusion in Independent Study Project (ISP) Collection by an authorized administrator of SIT Digital Collections. For more information, please contact [email protected]. 1 Large Mammals of the High Andes A Survey of Community Composition and Habitat Usage in the Highlands of Angochagua Fox, Sage Academic Director: Silva, Xavier, PhD Project Advisor: Laguna, Andres Oregon State University Fisheries and Wildlife Parroquia de Angochagua, Provincia de Imbabura, Ecuador Submitted in partial fulfillment of the requirements for Ecuador: Comparative Ecology and Conservation, SIT Study Abroad, Spring 2018 LARGE MAMMALS OF THE HIGH ANDES 2 Acknowledgements This project was made possible through continued support from the group Big Mammals Conservation. Andres Laguna, the executive director of the BMC, was responsible for all training and instruction during the project, and assisted with transect surveying, camera trap setup, analysis, and all logistics. -

ECUADOR 2012 NOVEMBER 13-December19 in 2011 When I
ECUADOR 2012 NOVEMBER 13-December19 In 2011 when I saw Wildwings offered a trip to see Spectacled bear and Mountain tapir in Ecuador, and that apparently the tapir was not too difficult I booked this tour. This tour was lead by Richard Webb and was mainly oriented to see these 2 species, but there was also a few days stay in Amazonia at Napo Wildlife center and short visits at Maqipucuna reserve, Cabanas San Isidro and Wildsumaco . InAmazonia we saw 3 Giant otter, 1 Collared peccary, Golden mantled tamarin, Pygmy marmoset, Common squirrel monkey, White fronted capuchin, Noisy night monkey, Double browed titi, Venezuelan red Howler monkey, 1 Spider monkey and 1 Brown-throated three-toed sloth, but we missed Equatorial saki and Common woolly monkey. All these species were in low numbers, but it is true that we didn't walk a lot. At Maqipucuna we didn't see any mammal. At Wild Sumaco we got a Red tailed squirrel and saw 10 of them at Cabanas San Isidro with 1 Black agouti and a Long-tailed weasel, plus footprints of Mountain tapir. For the bear and the tapir we tried several places, but mainly Papallacta in Coca Cayambe reserve. In 4 days we saw from the roads: 2 dfferents bears, a few White tailed deers, 1 Culpeo, 2 Forest rabbits but no tapir. By night several members of the group glimpsed a deer, either Little red brocket or Northern Pudu. Our local guide, Patricio Pillajo told me several times that to see the tapir from the roads was very rare. -

Hunting Trophies Guide D'identification CITES – Trophées De Chasse Guía De Identificación De CITES – Trofeos De Caza
CITES Identification Guide – Hunting Trophies Guide d'identification CITES – Trophées de chasse Guía de identificación de CITES – Trofeos de caza Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) CITES Identification Guide - Hunting Trophies Guide to the Identification of Game Mammals Controlled under the Convention on International Trade in Endangered Species of Wild Fauna and Flora Guide d'identification CITES - Trophées de chasse Guide d'identification des mammifères de chasse protégés par la Convention sur le commerce international des espèces de faune et de flore sauvages menacées d'extinction Guía de identificación de CITES - Trofeos de caza Guía de identificación de los mamiferos de caza protegidos por la Convención sobre el Comercio International de Especies Amenazadas de Fauna y Flora Silvestres An initiative of Environment Canada Une initiative d'Environnement Canada Una iniciativa del Ministerio del Medio Ambiente del Canadá Published by Authority of the Minister of Environment. © Minister of Supply and Services Canada, 2003. National Library of Canada cataloguing in publication data Main entry under title: CITES identification guide – hunting trophies : guide to the identification of game mammals controlled under the Convention on International Trade in Endangered Species of Wild Fauna and Flora = Guide d’identification CITES – trophées de chasse : guide d’identification des mammifères de chasse protégés par la Convention sur le commerce international des espèces de faune et de flore sauvages menacées d’extinction. = Guia de identificacion de CITES : trofeos de caza : guia de identificacion de los mamiferos de caza protegidos por la Convencion sobre el Comercio International de Especies Amenazadas de Fauna y Flora Silvestres Text in English, French and Spanish. -

Tapir Conservation
Volume 12 / Number 2 December 2003 Tapir Conservation The Newsletter of the IUCN/SSC Tapir Specialist Group www.tapirspecialistgroup.org Edited by Siân S. Waters and Stefan Seitz Contents n Letter from the Chair page 3 n Tapir Specialist Group Conservation Fund page 5 n TSG Committee Reports page 7 n The Malay Tapir Conservation Workshop page 8 n Current Project Updates page 11 n News from Captivity page 16 n Contributed Papers page 18 n Asking the Experts page 26 n Tapir Specialist Group Membership Directory page 30 Printing and distribution of the Tapir Conservation Newsletter See back cover and is supported by the Houston Zoo, page 2 for details. 1513 N. MacGregor, Houston, Texas 77030, United States, http://www.houstonzoo.org Tapir Conservation – The Newsletter of the IUCN/SSC Tapir Specialist Group Contents Volume 12, Number 2, December 2003 Abbreviation: Tapir Cons. 3 Letter from the Chair Editors Siân S. Waters – Contributions 5 TSG News CEI Consultancy Ltd., 14 Lindsay Garden 5 TSG Conservation Fund 2003 Tredegar, Gwent NP22 4RP, UK 7 TSG Committee Reports E-mail: [email protected] 7 Update from the TSG Veterinary Committee 7 Update from the TSG Zoo Committee Stefan Seitz – Layout & Graphics Bonndorfer Strasse 19 8 Conservation 68239 Mannheim, Germany 8 The Malay Tapir Conservation Workshop: A Major Success Phone & Fax: ++49 (0)621 47 14 28 E-mail: [email protected] 11 Current Project Updates Editorial Board Patrícia Medici – Chair, Tapir Specialist Group (TSG) 11 C o l o m b i a Avenida Perdizes, 285, Vila São Paulo, 11 Ecology and Conservation of the Mountain Tapir in the Teodoro Sampaio, São Paulo, Brazil 19280-000 Central Andes of Colombia Phone & Fax: ++55 (18) 3282 4690 13 Tolima Jimba Kush E-mail: [email protected] or [email protected] 14 E c u a d o r 14 Attitudes to Tapirs, Wilderness, and Wildlife Conservation Charles R. -

Official Measurer's Manual
Offi cial Measurer’s Manual Th e acceptance or denial of all entries are at the discretion of Safari Club International, its Board and committees . Entries are subject to review by the Record Book Committee of SCI at any time . Text by Jack Schwabland and L . Irvin Barnhart/Complex Antler Measuring Supplement contributed by Gabriel Paz and Gilbert Aguilar Revised September 2019 Table of Contents INTRODUCTION . 4 GENERAL INSTRUCTIONS FOR ALL ENTRIES Who may measure . 4 Who may enter . 4 What may be entered . 4 Where to enter . 5 When to enter . 5 How to enter . 6 When to measure . 6 Measuring tools, methods, fractions, etc . 6 Damaged and/or repaired trophies . 7 Sawn-apart or split skulls . 7 Carding off a broken horn, tusk or antler point . 7 Photographs . 8 Certification . 8 Remeasurement . 8 Major Awards and Alternative Method Awards . 9 Criteria for accepting animals . 9 Lawful and ethical hunting . 9 METHODS FOR MEASURING HORNS Method 1 For most animals with simple horns . 11 Method 2 For spiral-horned animals . 13 Method 3 For most wild cattle . 14 Method 4 For African buffalo . 15 Method 5 For common wildebeest and takin . 17 Method 6 For black wildebeest . 18 Method 7 For muskox . 19 Method 8 For rhinoceros . 20 Method 9 For multi-horned sheep . 21 Method 10 For pronghorn antelope . 22 Method 11 For wild sheep, bharal, aoudad and tur . 23 METHODS FOR MEASURING TUSKS Method 12 For hippopotamus, pigs, water deer and musk deer . 25 Method 13 For walrus . 26 Method 14 For elephant . 27 METHOD FOR MEASURING SKULLS Method 15 For carnivores, peccaries and water chevrotain . -

Donors to the SSC Conservation Communications Programme and the Deer Action Plan
Donors to the SSC Conservation Communications Programme and the Deer Action Plan The IUCN/Species Survival Commission is committed to communicate important species conservation information to natural resource managers, decision-makers and others whose actions affect the conservation of biodiversity. The SSC’s Action Plans, Occasional Papers, news magazine (Species), Membership Directory and other publications are supported by a wide variety of generous donors including: The Sultanate of Oman established the Peter Scott IUCN/SSC Action Plan Fund in 1990. The Fund supports Action Plan development and implementation; to date, more than 80 grants have been made from the Fund to Specialist Groups. As a result, the Action Plan Programme has progressed at an accelerated level and the network has grown and matured significantly. The SSC is grateful to the Sultanate of Oman for its confidence in and support for species conservation worldwide. The Chicago Zoological Society (CZS) provides significant in-kind and cash support to the SSC, including grants for special projects, editorial and design services, staff secondments and related support services. The mission of CZS is to help people develop a sustainable and harmonious relationship with nature. The Zoo carries out its mission by informing and inspiring 2,000,000 annual visitors, serving as a refuge for species threatened with extinction, developing scientific approaches to manage species successfully in zoos and the wild, and working with other zoos, agencies, and protected areas around the world to conserve habitats and wildlife. The Council of Agriculture (COA), Taiwan has awarded major grants to the SSC’s Wildlife Trade Programme and Conservation Communications Programme.