GAK (1C2): Sc-56903
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Supplementary Information Material and Methods
MCT-11-0474 BKM120: a potent and specific pan-PI3K inhibitor Supplementary Information Material and methods Chemicals The EGFR inhibitor NVP-AEE788 (Novartis), the Jak inhibitor I (Merck Calbiochem, #420099) and anisomycin (Alomone labs, # A-520) were prepared as 50 mM stock solutions in 100% DMSO. Doxorubicin (Adriablastin, Pfizer), EGF (Sigma Ref: E9644), PDGF (Sigma, Ref: P4306) and IL-4 (Sigma, Ref: I-4269) stock solutions were prepared as recommended by the manufacturer. For in vivo administration: Temodal (20 mg Temozolomide capsules, Essex Chemie AG, Luzern) was dissolved in 4 mL KZI/glucose (20/80, vol/vol); Taxotere was bought as 40 mg/mL solution (Sanofi Aventis, France), and prepared in KZI/glucose. Antibodies The primary antibodies used were as follows: anti-S473P-Akt (#9271), anti-T308P-Akt (#9276,), anti-S9P-GSK3β (#9336), anti-T389P-p70S6K (#9205), anti-YP/TP-Erk1/2 (#9101), anti-YP/TP-p38 (#9215), anti-YP/TP-JNK1/2 (#9101), anti-Y751P-PDGFR (#3161), anti- p21Cip1/Waf1 (#2946), anti-p27Kip1 (#2552) and anti-Ser15-p53 (#9284) antibodies were from Cell Signaling Technologies; anti-Akt (#05-591), anti-T32P-FKHRL1 (#06-952) and anti- PDGFR (#06-495) antibodies were from Upstate; anti-IGF-1R (#SC-713) and anti-EGFR (#SC-03) antibodies were from Santa Cruz; anti-GSK3α/β (#44610), anti-Y641P-Stat6 (#611566), anti-S1981P-ATM (#200-301), anti-T2609 DNA-PKcs (#GTX24194) and anti- 1 MCT-11-0474 BKM120: a potent and specific pan-PI3K inhibitor Y1316P-IGF-1R were from Bio-Source International, Becton-Dickinson, Rockland, GenTex and internal production, respectively. The 4G10 antibody was from Millipore (#05-321MG). -

Profiling Data
Compound Name DiscoveRx Gene Symbol Entrez Gene Percent Compound Symbol Control Concentration (nM) JNK-IN-8 AAK1 AAK1 69 1000 JNK-IN-8 ABL1(E255K)-phosphorylated ABL1 100 1000 JNK-IN-8 ABL1(F317I)-nonphosphorylated ABL1 87 1000 JNK-IN-8 ABL1(F317I)-phosphorylated ABL1 100 1000 JNK-IN-8 ABL1(F317L)-nonphosphorylated ABL1 65 1000 JNK-IN-8 ABL1(F317L)-phosphorylated ABL1 61 1000 JNK-IN-8 ABL1(H396P)-nonphosphorylated ABL1 42 1000 JNK-IN-8 ABL1(H396P)-phosphorylated ABL1 60 1000 JNK-IN-8 ABL1(M351T)-phosphorylated ABL1 81 1000 JNK-IN-8 ABL1(Q252H)-nonphosphorylated ABL1 100 1000 JNK-IN-8 ABL1(Q252H)-phosphorylated ABL1 56 1000 JNK-IN-8 ABL1(T315I)-nonphosphorylated ABL1 100 1000 JNK-IN-8 ABL1(T315I)-phosphorylated ABL1 92 1000 JNK-IN-8 ABL1(Y253F)-phosphorylated ABL1 71 1000 JNK-IN-8 ABL1-nonphosphorylated ABL1 97 1000 JNK-IN-8 ABL1-phosphorylated ABL1 100 1000 JNK-IN-8 ABL2 ABL2 97 1000 JNK-IN-8 ACVR1 ACVR1 100 1000 JNK-IN-8 ACVR1B ACVR1B 88 1000 JNK-IN-8 ACVR2A ACVR2A 100 1000 JNK-IN-8 ACVR2B ACVR2B 100 1000 JNK-IN-8 ACVRL1 ACVRL1 96 1000 JNK-IN-8 ADCK3 CABC1 100 1000 JNK-IN-8 ADCK4 ADCK4 93 1000 JNK-IN-8 AKT1 AKT1 100 1000 JNK-IN-8 AKT2 AKT2 100 1000 JNK-IN-8 AKT3 AKT3 100 1000 JNK-IN-8 ALK ALK 85 1000 JNK-IN-8 AMPK-alpha1 PRKAA1 100 1000 JNK-IN-8 AMPK-alpha2 PRKAA2 84 1000 JNK-IN-8 ANKK1 ANKK1 75 1000 JNK-IN-8 ARK5 NUAK1 100 1000 JNK-IN-8 ASK1 MAP3K5 100 1000 JNK-IN-8 ASK2 MAP3K6 93 1000 JNK-IN-8 AURKA AURKA 100 1000 JNK-IN-8 AURKA AURKA 84 1000 JNK-IN-8 AURKB AURKB 83 1000 JNK-IN-8 AURKB AURKB 96 1000 JNK-IN-8 AURKC AURKC 95 1000 JNK-IN-8 -

Application of a MYC Degradation
SCIENCE SIGNALING | RESEARCH ARTICLE CANCER Copyright © 2019 The Authors, some rights reserved; Application of a MYC degradation screen identifies exclusive licensee American Association sensitivity to CDK9 inhibitors in KRAS-mutant for the Advancement of Science. No claim pancreatic cancer to original U.S. Devon R. Blake1, Angelina V. Vaseva2, Richard G. Hodge2, McKenzie P. Kline3, Thomas S. K. Gilbert1,4, Government Works Vikas Tyagi5, Daowei Huang5, Gabrielle C. Whiten5, Jacob E. Larson5, Xiaodong Wang2,5, Kenneth H. Pearce5, Laura E. Herring1,4, Lee M. Graves1,2,4, Stephen V. Frye2,5, Michael J. Emanuele1,2, Adrienne D. Cox1,2,6, Channing J. Der1,2* Stabilization of the MYC oncoprotein by KRAS signaling critically promotes the growth of pancreatic ductal adeno- carcinoma (PDAC). Thus, understanding how MYC protein stability is regulated may lead to effective therapies. Here, we used a previously developed, flow cytometry–based assay that screened a library of >800 protein kinase inhibitors and identified compounds that promoted either the stability or degradation of MYC in a KRAS-mutant PDAC cell line. We validated compounds that stabilized or destabilized MYC and then focused on one compound, Downloaded from UNC10112785, that induced the substantial loss of MYC protein in both two-dimensional (2D) and 3D cell cultures. We determined that this compound is a potent CDK9 inhibitor with a previously uncharacterized scaffold, caused MYC loss through both transcriptional and posttranslational mechanisms, and suppresses PDAC anchorage- dependent and anchorage-independent growth. We discovered that CDK9 enhanced MYC protein stability 62 through a previously unknown, KRAS-independent mechanism involving direct phosphorylation of MYC at Ser . -

The Number of Genes
Table S1. The numbers of KD genes in each KD time The number The number The number The number Cell lines of genes of genes of genes of genes (96h) (120h) (144h) PC3 3980 3822 128 1725 A549 3724 3724 0 0 MCF7 3688 3471 0 1837 HT29 3665 3665 0 0 A375 3826 3826 0 0 HA1E 3801 3801 0 0 VCAP 4134 34 4121 0 HCC515 3522 3522 0 0 Table S2. The predicted results in the PC3 cell line on the LINCS II data id target rank A07563059 ADRB2 48 A12896037 ADRA2C 91 A13021932 YES1 77 PPM1B;PPP1CC;PPP2CA; A13254067 584;1326;297;171;3335 PTPN1;PPP2R5A A16347691 GMNN 2219 PIK3CB;MTOR;PIK3CA;PIK A28467416 18;10;9;13;8 3CG;PIK3CD A28545468 EHMT2;MAOB 14;67 A29520968 HSPB1 1770 A48881734 EZH2 1596 A52922642 CACNA1C 201 A64553394 ADRB2 155 A65730376 DOT1L 3764 A82035391 JUN 378 A82156122 DPP4 771 HRH1;HTR2C;CHRM3;CH A82772293 2756;2354;2808;2367 RM1 A86248581 CDA 1785 A92800748 TEK 459 A93093700 LMNA 1399 K00152668 RARB 105 K01577834 ADORA2A 525 K01674964 HRH1;BLM 31;1314 K02314383 AR 132 K03194791 PDE4D 30 K03390685 MAP2K1 77 K06762493 GMNN;APEX1 1523;2360 K07106112 ERBB4;ERBB2;EGFR 497;60;23 K07310275 AKT1;MTOR;PIK3CA 13;12;1 K07753030 RGS4;BLM 3736;3080 K08109215 BRD2;BRD3;BRD4 1413;2786;3 K08248804 XIAP 88 K08586861 TBXA2R;MBNL1 297;3428 K08832567 GMNN;CA12 2544;50 LMNA;NFKB1;APEX1;EH K08976401 1322;341;3206;123 MT2 K09372874 IMPDH2 232 K09711437 PLA2G2A 59 K10859802 GPR119 214 K11267252 RET;ALK 395;760 K12609457 LMNA 907 K13094524 BRD4 7 K13662825 CDK4;CDK9;CDK5;CDK1 34;58;13;18 K14704277 LMNA;BLM 1697;1238 K14870255 AXL 1696 K15170068 MAN2B1 1756 K15179879 -

Lipid Metabolic Reprogramming: Role in Melanoma Progression and Therapeutic Perspectives
cancers Review Lipid metabolic Reprogramming: Role in Melanoma Progression and Therapeutic Perspectives 1, 1, 1 2 1 Laurence Pellerin y, Lorry Carrié y , Carine Dufau , Laurence Nieto , Bruno Ségui , 1,3 1, , 1, , Thierry Levade , Joëlle Riond * z and Nathalie Andrieu-Abadie * z 1 Centre de Recherches en Cancérologie de Toulouse, Equipe Labellisée Fondation ARC, Université Fédérale de Toulouse Midi-Pyrénées, Université Toulouse III Paul-Sabatier, Inserm 1037, 2 avenue Hubert Curien, tgrCS 53717, 31037 Toulouse CEDEX 1, France; [email protected] (L.P.); [email protected] (L.C.); [email protected] (C.D.); [email protected] (B.S.); [email protected] (T.L.) 2 Institut de Pharmacologie et de Biologie Structurale, CNRS, Université Toulouse III Paul-Sabatier, UMR 5089, 205 Route de Narbonne, 31400 Toulouse, France; [email protected] 3 Laboratoire de Biochimie Métabolique, CHU Toulouse, 31059 Toulouse, France * Correspondence: [email protected] (J.R.); [email protected] (N.A.-A.); Tel.: +33-582-7416-20 (J.R.) These authors contributed equally to this work. y These authors jointly supervised this work. z Received: 15 September 2020; Accepted: 23 October 2020; Published: 27 October 2020 Simple Summary: Melanoma is a devastating skin cancer characterized by an impressive metabolic plasticity. Melanoma cells are able to adapt to the tumor microenvironment by using a variety of fuels that contribute to tumor growth and progression. In this review, the authors summarize the contribution of the lipid metabolic network in melanoma plasticity and aggressiveness, with a particular attention to specific lipid classes such as glycerophospholipids, sphingolipids, sterols and eicosanoids. -

1 Kinobead Profiling Reveals Reprogramming of B-Cell Receptor Signaling in Response to Therapy Within Primary CLL Cells. Linley
bioRxiv preprint doi: https://doi.org/10.1101/841312; this version posted November 14, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Kinobead profiling reveals reprogramming of B-cell receptor signaling in response to therapy within primary CLL cells. Linley AJ1, Griffin R2, Cicconi S2, D’Avola A3$, MacEwan DJ4, Pettit AR1, Kalakonda N1, Packham G3, Prior IA5, Slupsky JR1. 1. Department of Molecular and Clinical Cancer Medicine, Institute of Translational Medicine, University of Liverpool, Sherrington Building, Ashton Street, Liverpool, UK. 2. CRUK Clinical Trials Unit, University of Liverpool, Waterhouse Building, Ashton Street, Liverpool. 3. Southampton Cancer Research UK Centre, Cancer Sciences Unit, Faculty of Medicine, University of Southampton, Southampton, UK. 4. Department of Molecular and Clinical Pharmacology, Institute of Translational Medicine, University of Liverpool, Sherrington Building, Ashton Street, Liverpool, UK. 5. Department of Cellular and Molecular Physiology, Institute of Translational Medicine, University of Liverpool, Nuffield Wing, Crown Street, Liverpool, UK. $ Current Address: The Francis Crick Institute, 1 Midland Road, London, UK. Corresponding author: Dr Adam J Linley, Department of Molecular and Clinical Cancer Medicine, Institute of Translational Medicine, University of Liverpool, Sherrington Building, Ashton Street, Liverpool, UK; [email protected]; +44(0)151 794 5310 Running head: Therapy brings about BCR signal changes. Key points 1. sIgM signaling patterns alter following in vivo therapy using either chemoimmunotherapy or ibrutinib. 2. Kinobeads provide a novel method for high-resolution investigation of signaling in primary CLL cells. 1 bioRxiv preprint doi: https://doi.org/10.1101/841312; this version posted November 14, 2019. -

Inhibition of ERK 1/2 Kinases Prevents Tendon Matrix Breakdown Ulrich Blache1,2,3, Stefania L
www.nature.com/scientificreports OPEN Inhibition of ERK 1/2 kinases prevents tendon matrix breakdown Ulrich Blache1,2,3, Stefania L. Wunderli1,2,3, Amro A. Hussien1,2, Tino Stauber1,2, Gabriel Flückiger1,2, Maja Bollhalder1,2, Barbara Niederöst1,2, Sandro F. Fucentese1 & Jess G. Snedeker1,2* Tendon extracellular matrix (ECM) mechanical unloading results in tissue degradation and breakdown, with niche-dependent cellular stress directing proteolytic degradation of tendon. Here, we show that the extracellular-signal regulated kinase (ERK) pathway is central in tendon degradation of load-deprived tissue explants. We show that ERK 1/2 are highly phosphorylated in mechanically unloaded tendon fascicles in a vascular niche-dependent manner. Pharmacological inhibition of ERK 1/2 abolishes the induction of ECM catabolic gene expression (MMPs) and fully prevents loss of mechanical properties. Moreover, ERK 1/2 inhibition in unloaded tendon fascicles suppresses features of pathological tissue remodeling such as collagen type 3 matrix switch and the induction of the pro-fbrotic cytokine interleukin 11. This work demonstrates ERK signaling as a central checkpoint to trigger tendon matrix degradation and remodeling using load-deprived tissue explants. Tendon is a musculoskeletal tissue that transmits muscle force to bone. To accomplish its biomechanical function, tendon tissues adopt a specialized extracellular matrix (ECM) structure1. Te load-bearing tendon compart- ment consists of highly aligned collagen-rich fascicles that are interspersed with tendon stromal cells. Tendon is a mechanosensitive tissue whereby physiological mechanical loading is vital for maintaining tendon archi- tecture and homeostasis2. Mechanical unloading of the tissue, for instance following tendon rupture or more localized micro trauma, leads to proteolytic breakdown of the tissue with severe deterioration of both structural and mechanical properties3–5. -

(12) United States Patent (10) Patent No.: US 8,148,089 B2 Anderson Et Al
US008148089B2 (12) United States Patent (10) Patent No.: US 8,148,089 B2 Anderson et al. (45) Date of Patent: Apr. 3, 2012 (54) ALPHA-SYNUCLEIN KINASE FOREIGN PATENT DOCUMENTS (75) Inventors: John P. Anderson, San Francisco, CA WO WO 2006/073734 A2 T 2006 WO WO 2006/1248.92 A2 11/2006 (US); Kelly Banducci, Pleasanton, CA WO WO 2004/069175 A2 8, 2007 (US); Guriobal S. Basi, Palo Alto, CA WO WO 2007/089862 A2 8, 2007 (US); David Chereau, San Mateo, CA WO WO 2009/103010 A2 8, 2009 (US); Tamie J. Chilcote, San Francisco, CA (US); Normand L. Frigon, Jr., OTHER PUBLICATIONS Millbrae, CA (US); Jason Goldstein, U.S. Appl. No. 61/053,632, filed May 15, 2008, Anderson et al. Burlingame, CA (US); Irene Griswold, Anderson et al., “Phosphorylation of Ser-129 is the Dominant Patho San Francisco, CA (US) logical Modification of Alpha-Synclein in Familial and Sporadic Lewy Body Disease.” Journal of Biological Chemistry, 281 (73) Assignee: Elan Pharma International Limited, (40):29739-29752 (2006). Dublin (IE) Chen et al., “Alpha-Synuclein Phosphorylation Controls Neurotoxic ity and Inclusion Formation in a Drosophila Model of Parkinson (*) Notice: Subject to any disclaimer, the term of this Disease.” Nature Neuroscience, 8(5):657-663 (2005). patent is extended or adjusted under 35 Ellis et al., “Alpha-Synuclein is Phosphorylated by Members of the SCR Family of Protein-Tyrosine Kinases,” Journal of Biological U.S.C. 154(b) by 167 days. Chemistry, 276 (6):3879-3884 (2001). Inglis et al., “Polo-like kinase 2 (PLK2) phosphorylates alpha (21) Appl. -

Supplemental Materials: 1182 Supplemental Figures 1-7 1183 Supplemental Table 1 1184 Supplemental Movies 1-10 1185 1186
1181 Supplemental Materials: 1182 Supplemental Figures 1-7 1183 Supplemental Table 1 1184 Supplemental Movies 1-10 1185 1186 1187 1188 1189 Supplemental figure 1. Co-trafficking of HCV particles with host proteins. (A) 1190 Representative montages from live cell fluorescence microscopy of TC-core HCV particles co- 1191 trafficking with GFP-LC3. (B) Velocity of individual TC-core puncta co-trafficking with AP1 or 1192 AP2. Experiments were replicated twice. 1193 1194 1195 1196 1197 1198 1199 1200 1201 1202 1203 1204 Supplemental figure 2. 7745 Kd. Kd of compound 7745 for AAK1 as measured by a 1205 KINOMEscan kinase assay (DiscoverX). 50 1206 1207 1208 1209 Supplemental figure 3. Dose response of DENV infection to sunitinib and erlotinib. (A) 1210 Dose response of overall DENV infection to sunitinib and erlotinib measured by plaque assays at 1211 72 hours post-infection. Assay detection limit is marked with a dotted line. (B) Dose response of 1212 overall DENV infection to sunitinib/erlotinib combinations measured by luciferase assays at 48 1213 hours post-infection and normalized to DMSO control. All data shown are representative of at 1214 least two experiments with 6 biological replicates each. Shown are mean +/- s.d. PFU, plaque 1215 forming units. 1216 1217 1218 1219 1220 1221 1222 1223 Supplemental figure 4. Activity of sunitinib and erlotinib against additional flaviviruses. 1224 (A) Overall infection of murine embryonic fibroblast cells with WNV (black) and relative cell 1225 viability (blue) following 24 hour treatment with sunitinib measured by focus formation and 1226 MTT assays, respectively. (B) Overall infection of Huh7 cells with ZIKV (black) and relative 1227 cell viability (blue) following 48 hour treatment with sunitinib and erlotinib measured by plaque 1228 and alamarBlue assays, respectively. -

Evolution and Structure of Clinically Relevant Gene Fusions in Multiple Myeloma
Washington University School of Medicine Digital Commons@Becker Open Access Publications 2020 Evolution and structure of clinically relevant gene fusions in multiple myeloma Steven M. Foltz Qingsong Gao Christopher J. Yoon Hua Sun Lijun Yao See next page for additional authors Follow this and additional works at: https://digitalcommons.wustl.edu/open_access_pubs Authors Steven M. Foltz, Qingsong Gao, Christopher J. Yoon, Hua Sun, Lijun Yao, Yize Li, Reyka G. Jayasinghe, Song Cao, Justin King, Daniel R. Kohnen, Mark A. Fiala, Li Ding, and Ravi Vij ARTICLE https://doi.org/10.1038/s41467-020-16434-y OPEN Evolution and structure of clinically relevant gene fusions in multiple myeloma Steven M. Foltz 1,2, Qingsong Gao 1,2, Christopher J. Yoon1,2, Hua Sun1,2, Lijun Yao1,2, Yize Li1,2, ✉ ✉ Reyka G. Jayasinghe1,2, Song Cao1,2, Justin King1, Daniel R. Kohnen1, Mark A. Fiala1, Li Ding1,2,3,4 & Ravi Vij1,4 Multiple myeloma is a plasma cell blood cancer with frequent chromosomal translocations leading to gene fusions. To determine the clinical relevance of fusion events, we detect gene 1234567890():,; fusions from a cohort of 742 patients from the Multiple Myeloma Research Foundation CoMMpass Study. Patients with multiple clinic visits enable us to track tumor and fusion evolution, and cases with matching peripheral blood and bone marrow samples allow us to evaluate the concordance of fusion calls in patients with high tumor burden. We examine the joint upregulation of WHSC1 and FGFR3 in samples with t(4;14)-related fusions, and we illustrate a method for detecting fusions from single cell RNA-seq. -
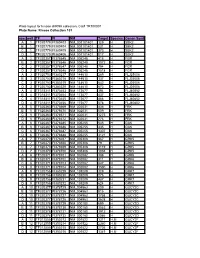
Kinase Collection 101 Row Well TF FI NM Target Location Spec
Plate layout for kinase shRNA collection, Cat# TR100001 Plate Name: Kinase Collection 101 row well TF FI NM Target LocationSpecies Genen Symbol A 1 TF320779 FI380403 NM_001101401 129 H SBK2 B 1 TF320779 FI380404 NM_001101401 231 H SBK2 C 1 TF320779 FI380405 NM_001101401 373 H SBK2 D 1 TF320779 FI380406 NM_001101401 817 H SBK2 A 2 TF320357 FI378645 NM_005248 415 H FGR B 2 TF320357 FI378646 NM_005248 1072 H FGR C 2 TF320357 FI378647 NM_005248 794 H FGR D 2 TF320357 FI378648 NM_005248 1518 H FGR A 3 TF320753 FI380217 NM_144610 289 H FLJ25006 B 3 TF320753 FI380218 NM_144610 137 H FLJ25006 C 3 TF320753 FI380219 NM_144610 642 H FLJ25006 D 3 TF320753 FI380220 NM_144610 673 H FLJ25006 A 4 TF318311 FI370453 NM_173677 394 H FLJ40852 B 4 TF318311 FI370454 NM_173677 437 H FLJ40852 C 4 TF318311 FI370455 NM_173677 466 H FLJ40852 D 4 TF318311 FI370456 NM_173677 578 H FLJ40852 A 5 TF320363 FI378669 NM_002031 436 H FRK B 5 TF320363 FI378670 NM_002031 509 H FRK C 5 TF320363 FI378671 NM_002031 1275 H FRK D 5 TF320363 FI378672 NM_002031 776 H FRK A 6 TF320367 FI378685 NM_005255 545 H GAK B 6 TF320367 FI378686 NM_005255 435 H GAK C 6 TF320367 FI378687 NM_005255 1307 H GAK D 6 TF320367 FI378688 NM_005255 3121 H GAK A 7 TF320370 FI378697 NM_005308 367 H GRK5 B 7 TF320370 FI378698 NM_005308 79 H GRK5 C 7 TF320370 FI378699 NM_005308 1119 H GRK5 D 7 TF320370 FI378700 NM_005308 1038 H GRK5 A 8 TF320371 FI378701 NM_002082 388 H GRK6 B 8 TF320371 FI378702 NM_002082 311 H GRK6 C 8 TF320371 FI378703 NM_002082 847 H GRK6 D 8 TF320371 FI378704 NM_002082 1590 H GRK6 A -
Live Imaging Screen Reveals That TYRO3 and GAK Ensure Accurate Spindle Positioning in Human Cells
ARTICLE https://doi.org/10.1038/s41467-019-10446-z OPEN Live imaging screen reveals that TYRO3 and GAK ensure accurate spindle positioning in human cells Benita Wolf1, Coralie Busso1 & Pierre Gönczy1 Proper spindle positioning is crucial for spatial cell division control. Spindle positioning in human cells relies on a ternary complex comprising Gαi1–3, LGN and NuMA, which anchors dynein at the cell cortex, thus enabling pulling forces to be exerted on astral microtubules. fi 1234567890():,; We develop a live imaging siRNA-based screen using stereotyped bronectin micropatterns to uncover components modulating spindle positioning in human cells, testing 1280 genes, including all kinases and phosphatases. We thus discover 16 components whose inactivation dramatically perturbs spindle positioning, including tyrosine receptor kinase 3 (TYRO3) and cyclin G associated kinase (GAK). TYRO3 depletion results in excess NuMA and dynein at the cortex during metaphase, similar to the effect of blocking the TYRO3 downstream tar- get phosphatidylinositol 3-kinase (PI3K). Furthermore, depletion of GAK leads to impaired astral microtubules, similar to the effect of downregulating the GAK-interactor Clathrin. Overall, our work uncovers components and mechanisms governing spindle positioning in human cells. 1 Swiss Institute for Experimental Cancer Research (ISREC), School of Life Sciences, Swiss Federal Institute of Technology Lausanne (EPFL), 1015 Lausanne, Switzerland. Correspondence and requests for materials should be addressed to P.G. (email: pierre.gonczy@epfl.ch) NATURE COMMUNICATIONS | (2019) 10:2859 | https://doi.org/10.1038/s41467-019-10446-z | www.nature.com/naturecommunications 1 ARTICLE NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-019-10446-z patial control of cell division is crucial during development network24.