Pb-651 PC Srivastava1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Two LITTLE KNOWN SPECIES of SPHENOPHYLLUM from the STEPHANIAN of SPAIN and FRANCE
Two LITTLE KNOWN SPECIES OF SPHENOPHYLLUM FROM THE STEPHANIAN OF SPAIN AND FRANCE ROBERT H. WAGNER Geology Department, University of Sheffield, England, U.K. ABSTRACT The impression species Sphenophyllum incisum 'Wagner is fully described for the first time. with due reference to intra-specific variation as shown by abundant mate• rial from a single locality in the Stephanian B of North-west Spain. The associated stro• bilus is also recorded. Sphenophyllum nageli Grand'Eury, a middle to late Canta• brian species, is redescribed from the type region in the Cevennes, southern France. Similar material is illustrated from Cantabrian and Stephanian A localities in North-west Spain. Brief comments are made on several other species of Sphenophyllum from the Stephanian of western Europe, and a range chart is provided. INTRODUCTION basis of a large collection of specimens from a single locality. Another little known species, Spheno• Doubingeris well overand twentyVetter (1954)years agopublishedsince phyllum nageli Grand'Eury, was actually ITa brief not~ on the various species of described in 1890, but hardly mentioned Sphenophyll1&m recorded from Stephanian afterwards. Doubinger and Vetter ignored strata. A monographic paper by Abbott it entirely in their synthesis of 1954, and (1958) filled in gaps with regard to the the species did not reappear in the literature American species; a brief note by Obrhel until 1970 when it Was mentioned again (1967) brought the description of Spheno• from the original type area, the Cevennes, phyllum nemejci Obrhel from the Stephanian by Bouroz, Gras and Wagner. The material of Bohemia; and recent work on the reported in that paper includes several Stephanian floras of Portugal and Spain well-preserved specimens which allow a has added several new species such as redescription of Sphenophyllum nageli Grand• Sphenophyllum guerreiroi Teixeira, Spheno• 'Eury, as provided in the present paper. -

Sementes Do Gênero Samaropsis Goeppert No Permiano Inferior Da Bacia Do Paraná, Sul Do Brasil
Rev. bras. paleontol. 10(2):95-106, Maio/Agosto 2007 © 2007 by the Sociedade Brasileira de Paleontologia SEMENTES DO GÊNERO SAMAROPSIS GOEPPERT NO PERMIANO INFERIOR DA BACIA DO PARANÁ, SUL DO BRASIL JULIANE MARQUES DE SOUZA & ROBERTO IANNUZZI Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, UFRGS, Cx. P. 15001, 91501-970, Porto Alegre, RS, Brasil. [email protected], [email protected] RESUMO – O afloramento Morro do Papaléo, localizado no município de Mariana Pimentel, Rio Grande do Sul, tem se mostrado uma importante fonte de sementes fósseis preservadas na forma de impressões, as quais são provenientes do Grupo Itararé e da Formação Rio Bonito, Permiano Inferior da bacia do Paraná. Neste estudo são descritos seis diferentes morfotipos para o gênero Samaropsis Goeppert que, possivelmente, correspondem a seis diferentes morfoespécies. Características como a largura da sarcotesta, a forma geral da semente e do nucelo e a textura de sua superfície permitiram a proposição de uma nova espécie, Samaropsis gigas nov. sp., e identificar a presença de Samaropsis kurtzii Leguizamón, S. aff. S. millaniana Oliveira & Pontes e S. aff. S. rigbyi Millan. Além disso, uns poucos espécimes foram classificados apenas a nível genérico, tendo sido designados como Samaropsis sp. 1 e Samaropsis sp. 2. Palavras-chave: Sementes fósseis, Grupo Itararé, Formação Rio Bonito, bacia do Paraná, Rio Grande do Sul. ABSTRACT – SEEDS OF THE GENUS SAMAROPSIS GOEPPERT IN THE LOWER PERMIAN OF THE PARANÁ BASIN, SOUTHERN BRAZIL. The Morro do Papaléo outcrop is located in Mariana Pimentel town, Rio Grande do Sul, and has been an important source of fossil seeds. -

Subclase Equisetidae ¿Tienes Alguna Duda, Sugerencia O Corrección Acerca De Este Taxón? Envíanosla Y Con Gusto La Atenderemos
subclase Equisetidae ¿Tienes alguna duda, sugerencia o corrección acerca de este taxón? Envíanosla y con gusto la atenderemos. Ver todas las fotos etiquetadas con Equisetidae en Banco de Imagénes » Descripción de WIKIPEDIAES Ver en Wikipedia (español) → Ver Pteridophyta para una introducción a las plantas Equisetos vasculares sin semilla Rango temporal: Devónico-Holoceno PreЄ Є O S D C P T J K Pg N Los equisetos , llamados Equisetidae en la moderna clasificación de Christenhusz et al. 2011,[1] [2] [3] o también Equisetopsida o Equisetophyta, y en paleobotánica es más común Sphenopsida, son plantas vasculares afines a los helechos que aparecieron en el Devónico, pero que actualmente sobrevive únicamente el género Equisetum, si bien hay representantes de órdenes extintos que se verán en este artículo. Este grupo es monofilético, aun con sus representantes extintos, debido a su morfología distintiva. Son plantas pequeñas, aunque en el pasado una variedad de calamitácea alcanzó los 15 metros durante el pérmico.[4] Índice 1 Filogenia 1.1 Ecología y evolución 2 Taxonomía 2.1 Sinonimia Variedades de Equisetum 2.2 Sistema de Christenhusz et al. 2011 Taxonomía 2.3 Clasificación sensu Smith et al. 2006 2.4 Otras clasificaciones Reino: Plantae 3 Caracteres Viridiplantae 4 Véase también Streptophyta 5 Referencias Streptophytina 6 Bibliografía Embryophyta (sin rango) 7 Enlaces externos Tracheophyta Euphyllophyta Monilophyta Filogenia[editar] Equisetopsida o Sphenopsida Introducción teórica en Filogenia Clase: C.Agardh 1825 / Engler 1924 Equisetidae Los análisis moleculares y genéticos de filogenia solo Subclase: se pueden hacer sobre representantes vivientes, Warm. 1883 como circunscripto según Smith et al. (2006) (ver la Órdenes ficha), al menos Equisetales es monofilético (Pryer et Equisetales (DC. -

Permophiles Issue #28 1996 1
Permophiles Issue #28 1996 1. SECRETARY’S NOTE I should like to thank all those who contributed to this issue of I have discussed the matter of financial support for Permophiles @Permophiles@. The next issue will be in November 1996 and with many of the members of the Permian Subcommission; the will be prepared by the new secretary, CIaude Spinosa. Please consensus is that we should request voluntary donations to offset send your contributions to him (see note below from Forthcoming processing, mailing and paper costs. We are asking for contribu- Secretary). tions of $10 to $25. However, Permophiles will be mailed to ev- I should like to express my gratitude to those of you who have eryone on the mailing list regardless of whether a contribution is submitted contributions during my eight year term of office. You made. have helped make =Permophiles@ a useful, informative, and timely We have established access to Permophiles via the Internet; the Newsletter of the Subcommission on Permian Stratigraphy. address will be: J. Utting http://earth.idbsu.edu/permian/permophiles Geological Survey of Canada (Calgary) 3303 - 33rd Street N.W. Our intention is to provide a version of Permophiles that is readily Calgary, Alberta, Canada T2L 2A7 available through the Internet. The Internet version of Permophiles Phone (404) 292-7093 FAX (403) 292-6014 will have multiple formats. Initially the format of the Internet ver- E-mail INTERNET address: [email protected] sion will be different from the official paper version. Because new taxonomic names can not be published in Permophiles, a hard copy, downloaded from the interned will suffice for many of us. -

Bioestratigrafía Del Paleozoico Superior De América Del Sur: Primera Etapa De Trabajo Hacia Una Nueva Propuesta Cronoestratigr
Asociación Geológica Argentina Serie D: Publicación especial Nº 11 Bioestratigrafía del Paleozoico Superior de América del Sur: Primera Etapa de Trabajo Hacia una Nueva Propuesta Cronoestratigráfica La impresión de esta publicación fue posible por la colaboración de: la Agencia Nacional de Promoción Científica y Tecnológica a través del PICT-R 00313/1-2 Foto de tapa: Hugo Carrizo. Extremo Norte de la Sierra de Narváez (Sistema de Famatina). Vista hacia el norte del río Chaschuil. Formación De La Cuesta (Pérmico).Sección Superior. Sedimentitas fluviales con estratificación tangencial. Diseño y composición: M. Laura Iribas. Correo: [email protected] Se terminó de imprimir en el mes de septiembre de 2007 en los talleres del Instituto Salesiano de Artes Gráficas, Don Bosco 4053, Buenos Aires, Argentina. ISSN 0328-2767. Reservados todos los derechos. BIOESTRATIGRAFÍA DEL PALEOZOICO SUPERIOR DE AMÉRICA DEL SUR: PRIMERA ETAPA DE TRABAJO HACIA UNA NUEVA PROPUESTA CRONOESTRATIGRÁFICA Convocada por: Paleontólogos de la UBA y de la Fundación Lillo. Auspiciada por: Comité Organizador de la XI Reunião de PaleobotânicosPaleobotânicos y por el Comité Organizador del XIII Simposio Argentino de Paleobotánica y Palinología (Bahía Blanca) Argentina 2006. Comité organizador local Dr. Carlos L. Azcuy CONTENIDO Prólogo .......................................................................................................................................................................................... 5 Introducción ............................................................................................................................................................................. -

A Reappraisal of Mississippian (Tournaisian and Visean) Adpression Floras from Central and Northwestern Europe
Zitteliana A 54 (2014) 39 A reappraisal of Mississippian (Tournaisian and Visean) adpression floras from central and northwestern Europe Maren Hübers1, Benjamin Bomfleur2*, Michael Krings3, Christian Pott2 & Hans Kerp1 1Forschungsstelle für Paläobotanik am Geologisch-Paläontologischen Institut, Westfälische Zitteliana A 54, 39 – 52 Wilhelms-Universität Münster, Heisenbergstraße 2, 48149 Münster, Germany München, 31.12.2014 2Swedish Museum of Natural History, Department of Paleobiology, Box 50007, SE-104 05, Stockholm, Sweden Manuscript received 3Department für Geo- und Umweltwissenschaften, Paläontologie und Geobiologie, Ludwig- 01.02.2014; revision Maximilians-Universität, and Bayerische Staatssammlung für Paläontologie und Geologie, Richard- accepted 07.04.2014 Wagner-Straße 10, 80333 Munich, Germany ISSN 1612 - 412X *Author for correspondence and reprint requests: E-mail: [email protected] Abstract Mississippian plant fossils are generally rare, and in central and northwestern Europe especially Tournaisian to middle Visean fossil floras are restricted to isolated occurrences. While sphenophytes and lycophytes generally are represented by only a few widespread and long-ranging taxa such as Archaeocalamites radiatus, Sphenophyllum tenerrimum and several species of Lepidodendropsis and Lepido- dendron, Visean floras in particular show a remarkably high diversity of fern-like foliage, including filiform types (Rhodea, Diplotmema), forms with bipartite fronds (Sphenopteridium, Diplopteridium, Spathulopteris, Archaeopteridium), others with monopodial, pinnate fronds (Anisopteris, Fryopsis) and still others characterized by several-times pinnate fronds (e.g., Adiantites, Triphyllopteris, Sphenopteris, Neu- ropteris). Most of these leaf types have been interpreted as belonging to early seed ferns, whereas true ferns seem to have been rare or lacking in impression/compression floras. In the upper Visean, two types of plant assemblages can be distinguished, i.e., the northern Kohlenkalk-type and the south-eastern Kulm-type assemblage. -

Three New Fern Fronds from the Glossopteris Flora of India
THREE NEW FERN FRONDS FROM THE GLOSSOPTERIS FLORA OF INDIA P. K. MAITHY Birbal Sahni Institute of Palaeobotany, Lucknow-226007 ABSTRACT of dates, Damudopteris becomes synonym A new species of Neomariopteris and a new to Neomariopteris, because the former form species of Dichotomopteris is recorded. In addition has been published one month later. to this a new genus Santhalea is instituted. INTRODUCTION Neomariopteris khanU sp. novo Diagnosis - Fronds large, at least tri• the ferns from the Lower Gondwanas knowledge on the morphology of pinnate; catadromic, rachis winged, secon• OURof India has been advanced consider• dary rachis broad, emerge alternately at an ably from the recent work of Maithy (1974a, angle of ± 60°; pinnae lanceolate; attached 1974b, 1975), Pant and Misra (1976) and alternate, sub-opposite or opposite from Pant and Khare (1974). Recently Maithy secondary rachis; lateral pinnules ovate, has revised the Lower Gondwana ferns from 1'0 cm long and 0'4 mm broad at base, i.e. India. On the basis of his revision, he has the length and breadth ratio of the pinnules instituted two new genera Neomariopteris is 2·5: I, lateral pinnules alternately and Dichotomopteris and an unrecorded form arranged, standing at right angles to rachis, Dizeugotheca. Pant and Khare (1974) and decurrent, attached by broad bases, lateral Pant and Misra (1976) have reported two fusion of two pinnules margin is ± 1/4 length new genera Damudopteris and Asansolia from of the pinnules from the base; apex acute; the Raniganj Coalfield. margin entire; both the margins show out• The present paper deals with three new ward curvature; terminal pinnules smaller fern fronds collected recently from than lateral pinnules, triangular in shape. -

Iop Newsletter 120
IOP NEWSLETTER 120 October 2019 CONTENTS Letter from the president Elections of IOP Executive Committee 2020 – Call for nominations Special issue in occasion of the 65th birthday of Hans Kerp Collection Spotlight: Cleveland Museum of Natural History Reflections from the ‘Earth Day’ 2019 Upcoming meetings IOP Logo: The evolution of plant architecture (© by A. R. Hemsley) 1 Letter from the president Greetings Members, This past quarter we have welcomed the publication Festschrifts celebrating our colleagues Hans Kerp (PalZ Paläontologische Zeitschrift, see below) and Gar Rothwell (International Journal of Plant Sciences v 180 nos. 7, 8). Thank you to the teams of editors and authors who have contributed to these issues illustrating the continuing strength of our discipline— and congratulations to Hans and Gar for inspiring such productivity by your admiring colleagues! Already three years have passed since our gathering for IOPC in Salvador, Brazil in 2016, and now planning is in full swing for next year’s meeting in Prague, 12-19 September 2020. This will be the 11th quadrennial meeting of IOP, held concurrently with the 15th International Palynological Conference. Please note the formal announcement in this newsletter (page 12, herein). Nominations for colleagues deserving of honorary membership are welcome at any time. As the submission of abstracts for IOPC/IPC presentations will be possible soon, we kindly remind the IOP Student Travel Awards. We will financially support about 5 to 7 PhD or MSc students in order to enable them to participate in the conference and present their research results in a talk. Recent PhD graduates will also qualify for these awards, if their completion was less than nine months prior to the time of the conference. -

Bioestratigrafía Del Pérmico De La Sierra De Los Llanos, La Rioja (Cuenca Paganzo), Sobre La Base De La Megaflora Y Su Correlación Con Áreas Relacionadas
Bioestratigrafía del Pérmico de la Sierra de Los Llanos, La Rioja (Cuenca Paganzo), sobre la base de la megaflora y su correlación con áreas relacionadas Eliana Paula Coturel Tesis para optar por el título de Doctor en Ciencias Naturales Director : Dr. Pedro Raúl Gutiérrez Codirector : Dr. Eduardo Morel Facultad de Ciencias Naturales y Museo Universidad Nacional de La Plata - 2013 - ELIANA PAULA COTUREL – BIOESTRATIGRAFÍA DEL PÉRMICO DE LA SIERRA DE LOS LLANOS … Quiero dedicar esta Tesis a mi mamá, Mónica Edith Emmerich. Creo que no hace falta dar razones de por qué una le dedica su Tesis a su Madre. Tampoco hay, en el rico idioma castellano, palabras suficientes para hacerlo. Agradezco a todos los que hicieron posible el desarrollo de esta Tesis. A mis directores, los Dres. Pedro R. Gutiérrez y Eduardo M. Morel, por la confianza depositada y por permitirme trabajar y aprender bajo su ala. A mis compañeras de equipo, Lucía Balarino y Bárbara Cariglino, por la ayuda y el apoyo que me dieron en todos estos años (nota aquí: Bárbara escaneó las láminas de Glossopteris que usé en el capítulo de morfografía, es importante mencionarlo). Las quiero mucho. Y a Gustavo Correa, por el apoyo en el viaje de campo y el tiempo que compartimos en el Museo. Lo normal sería ver aquí un listado de gente y miles de agradecimientos personales, pero a mí no me parece que sea el lugar (principalmente porque seguro que me olvido de alguien y queda muy feo al estar escrito!!). A cada persona que quiero, que aprecio, se lo hago saber (quizás no con palabras, pero sí con los gestos). -

Transformative Paleobotany
Chapter 6 Lower Permian Flora of the Sanzenbacher Ranch, Clay County, Texas William A. DiMichele1, Robert W. Hook2, Hans Kerp3, Carol L. Hotton1,4, Cindy V. Looy5 and Dan S. Chaney1 1NMNH Smithsonian Institution, Washington, DC, United States; 2The University of Texas at Austin, Austin, TX, United States; 3Westfälische Wilhelms-Universität Münster, Münster, Germany; 4National Institutes of Health, Bethesda, MD, United States; 5University of California Berkeley, Berkeley, CA, United States 1. INTRODUCTION 1985; Broutin, 1986; Popa, 1999; Steyer et al., 2000; Wagner and Mayoral, 2007; Bercovici and Broutin, 2008; Since 1989, field parties supported by the U.S. National Barthel, 2009; Wagner and Álvarez-Vázquez, 2010; Museum of Natural History have obtained large collections Barthel and Brauner, 2015). Furthermore, because this of mainly Permian plant fossils from north central Texas. locality was collected on three occasions over a time period This work was undertaken to study known localities and to of 50 years and by different parties, comparative analysis of find new fossiliferous deposits that would contribute to a the Sanzenbacher collections provides a basis for assessing better understanding of floral and paleoenvironmental sites that have comparable histories. changes within the region during the early Permian. From the outset, the effort was interdisciplinary and grew, through the contributions of nearly 20 paleobotanists, 2. GEOLOGY palynologists, invertebrate and vertebrate paleontologists, Clay County is the only county in the Permo-Carboniferous and sedimentary geologists of several subdisciplines, to be outcrop belt of north central Texas that lacks marine rocks. quite comprehensive. Our reporting of results, however, has These alluvial sediments accumulated east of a broad been influenced by unexpected developments, including the coastal plain that bordered the Eastern Shelf of the Midland discovery of new plant-fossil assemblages in areas once Basin. -
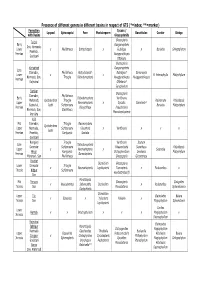
Presence of Different Genera in Different Basins in Respect of GTS
Presence of different genera in different basins in respect of GTS (*=index; **=marker) Formation Cycads / Lyopod Sphenopsid Fern Pteridosperm Benettiales Conifer Ginkgo with Basins Glossopterids Glossopteris Talchir Early Gangamopteris Son, Narmada, Lower x Phyllotheca Botrychiopsis x Rubidgea x Buriadia Ginkgophyton Pranhita, Permian Noeggerothiopis Godabari Ottokaria Glossopteris Karharbari Gangamopteris Late Damodar, Phyllotheca Botrychiopsis* Rubidgea* Samaropsis Lower x x B. heterophylla Platyphyllum Narmada, Son, Trizygia Dichotomopteris Noeggerothiopis Noeggarothiopsis Permian Rajmahal Ottokaria* Euryphyllum Barakar Glossopteris Damodar, Phyllotheca Early Dichotomopteris Vertibraria Mahanadi, Cyclodendron Trizygia Walkomella Rhipidopsis Upper Neomariopteris x Cycads: Barakaria* Rajmahal, lesbii Schizoneura Buriadia Platyphyllum Permian Dizagotheca Pseudocteris Narmada, Son, Stellotheca Macrotaeniopteris Pranhita Kulti Mid Damodar, Trizygia Neomaripteris Cyclodendron Upper Narmada, Schizoneura Gleopteris x Vertibraria x x x lesbii Permian Pranhita, Raniganjia Santalia Godabari Raniganj Trizygia Vertibraria Scutum Late Dichotomopteris Damodar Schizoneura Palaeovittaria Senotheca Rhipidopsis Upper x Neomariopteris x Searsolia Hingir Raniganjia Dictyopteridium Denkania Platyphyllum Permain Damodopteris Mahanadi, Son Phyllotheca Glossopteris Glossotheca Panchet Dicroidium Glossopteris Lower Damodar Trizygia x Neomariopteris Lepidopteris Taenopteris x Podozanites x Triassic Nidpur Schizoneura Kendostrobus(f) Son Marattiopsis Mid Parsora -

Diversity and Evolution of the Megaphyll in Euphyllophytes
G Model PALEVO-665; No. of Pages 16 ARTICLE IN PRESS C. R. Palevol xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect Comptes Rendus Palevol w ww.sciencedirect.com General palaeontology, systematics and evolution (Palaeobotany) Diversity and evolution of the megaphyll in Euphyllophytes: Phylogenetic hypotheses and the problem of foliar organ definition Diversité et évolution de la mégaphylle chez les Euphyllophytes : hypothèses phylogénétiques et le problème de la définition de l’organe foliaire ∗ Adèle Corvez , Véronique Barriel , Jean-Yves Dubuisson UMR 7207 CNRS-MNHN-UPMC, centre de recherches en paléobiodiversité et paléoenvironnements, 57, rue Cuvier, CP 48, 75005 Paris, France a r t i c l e i n f o a b s t r a c t Article history: Recent paleobotanical studies suggest that megaphylls evolved several times in land plant st Received 1 February 2012 evolution, implying that behind the single word “megaphyll” are hidden very differ- Accepted after revision 23 May 2012 ent notions and concepts. We therefore review current knowledge about diverse foliar Available online xxx organs and related characters observed in fossil and living plants, using one phylogenetic hypothesis to infer their origins and evolution. Four foliar organs and one lateral axis are Presented by Philippe Taquet described in detail and differ by the different combination of four main characters: lateral organ symmetry, abdaxity, planation and webbing. Phylogenetic analyses show that the Keywords: “true” megaphyll appeared at least twice in Euphyllophytes, and that the history of the Euphyllophytes Megaphyll four main characters is different in each case. The current definition of the megaphyll is questioned; we propose a clear and accurate terminology in order to remove ambiguities Bilateral symmetry Abdaxity of the current vocabulary.