Three New Fern Fronds from the Glossopteris Flora of India
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sementes Do Gênero Samaropsis Goeppert No Permiano Inferior Da Bacia Do Paraná, Sul Do Brasil
Rev. bras. paleontol. 10(2):95-106, Maio/Agosto 2007 © 2007 by the Sociedade Brasileira de Paleontologia SEMENTES DO GÊNERO SAMAROPSIS GOEPPERT NO PERMIANO INFERIOR DA BACIA DO PARANÁ, SUL DO BRASIL JULIANE MARQUES DE SOUZA & ROBERTO IANNUZZI Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, UFRGS, Cx. P. 15001, 91501-970, Porto Alegre, RS, Brasil. [email protected], [email protected] RESUMO – O afloramento Morro do Papaléo, localizado no município de Mariana Pimentel, Rio Grande do Sul, tem se mostrado uma importante fonte de sementes fósseis preservadas na forma de impressões, as quais são provenientes do Grupo Itararé e da Formação Rio Bonito, Permiano Inferior da bacia do Paraná. Neste estudo são descritos seis diferentes morfotipos para o gênero Samaropsis Goeppert que, possivelmente, correspondem a seis diferentes morfoespécies. Características como a largura da sarcotesta, a forma geral da semente e do nucelo e a textura de sua superfície permitiram a proposição de uma nova espécie, Samaropsis gigas nov. sp., e identificar a presença de Samaropsis kurtzii Leguizamón, S. aff. S. millaniana Oliveira & Pontes e S. aff. S. rigbyi Millan. Além disso, uns poucos espécimes foram classificados apenas a nível genérico, tendo sido designados como Samaropsis sp. 1 e Samaropsis sp. 2. Palavras-chave: Sementes fósseis, Grupo Itararé, Formação Rio Bonito, bacia do Paraná, Rio Grande do Sul. ABSTRACT – SEEDS OF THE GENUS SAMAROPSIS GOEPPERT IN THE LOWER PERMIAN OF THE PARANÁ BASIN, SOUTHERN BRAZIL. The Morro do Papaléo outcrop is located in Mariana Pimentel town, Rio Grande do Sul, and has been an important source of fossil seeds. -

Plant Fossils and Gondwana Flora
UNIT 12 PLANT FOSSILS AND GONDWANA FLORA Structure_____________________________________________________ 12.1 Introduction Vertebraria Expected Learning Outcomes Thinnfeldia 12.2 Plant Fossils Sigillaria Definition Nilssonia Classification Williamsonia Modes of Preservation Ptilophyllum Significance 12.5 Activity 12.3 Gondwana Flora of India 12.6 Summary 12.4 Descriptions of some Plant 12.7 Terminal Questions Fossils 12.8 References Glossopteris 12.9 Further/Suggested Readings Gangamopteris 12.10 Answers 12.1 INTRODUCTION The animals, plants and micro-organisms are the three main life forms surviving today. Even their fossilised remains are found in rocks that tell us about their past history. The animals comprise invertebrates and vertebrates. In Block 4, you will read about the invertebrates and their geological history that began in the latest Precambrian time. You also read about the microfossils in Unit 10 that too have a long geological record beginning from Precambrian onwards. In Unit 11, you read the evolutionary history of one of the vertebrate groups i.e., horse. In this unit, you will read the plant fossils and the Gondwana flora of India. Introduction to Palaeontology Block……………………………………………………………………………………………….….............….…........ 3 Like the kingdom Animalia, plants also form a separate kingdom known as the Plantae. It is thought that plants appeared first in the Precambrian, but their fossil record is poor. It is also proposed that earliest plants were aquatic and during the Ordovician period a transition from water to land took place that gave rise to non-vascular land plants. However, it was during the Silurian period, that the vascular plants appeared first on the land. The flowering plants emerged rather recently, during the Cretaceous period. -

Permophiles Issue #28 1996 1
Permophiles Issue #28 1996 1. SECRETARY’S NOTE I should like to thank all those who contributed to this issue of I have discussed the matter of financial support for Permophiles @Permophiles@. The next issue will be in November 1996 and with many of the members of the Permian Subcommission; the will be prepared by the new secretary, CIaude Spinosa. Please consensus is that we should request voluntary donations to offset send your contributions to him (see note below from Forthcoming processing, mailing and paper costs. We are asking for contribu- Secretary). tions of $10 to $25. However, Permophiles will be mailed to ev- I should like to express my gratitude to those of you who have eryone on the mailing list regardless of whether a contribution is submitted contributions during my eight year term of office. You made. have helped make =Permophiles@ a useful, informative, and timely We have established access to Permophiles via the Internet; the Newsletter of the Subcommission on Permian Stratigraphy. address will be: J. Utting http://earth.idbsu.edu/permian/permophiles Geological Survey of Canada (Calgary) 3303 - 33rd Street N.W. Our intention is to provide a version of Permophiles that is readily Calgary, Alberta, Canada T2L 2A7 available through the Internet. The Internet version of Permophiles Phone (404) 292-7093 FAX (403) 292-6014 will have multiple formats. Initially the format of the Internet ver- E-mail INTERNET address: [email protected] sion will be different from the official paper version. Because new taxonomic names can not be published in Permophiles, a hard copy, downloaded from the interned will suffice for many of us. -

Bioestratigrafía Del Paleozoico Superior De América Del Sur: Primera Etapa De Trabajo Hacia Una Nueva Propuesta Cronoestratigr
Asociación Geológica Argentina Serie D: Publicación especial Nº 11 Bioestratigrafía del Paleozoico Superior de América del Sur: Primera Etapa de Trabajo Hacia una Nueva Propuesta Cronoestratigráfica La impresión de esta publicación fue posible por la colaboración de: la Agencia Nacional de Promoción Científica y Tecnológica a través del PICT-R 00313/1-2 Foto de tapa: Hugo Carrizo. Extremo Norte de la Sierra de Narváez (Sistema de Famatina). Vista hacia el norte del río Chaschuil. Formación De La Cuesta (Pérmico).Sección Superior. Sedimentitas fluviales con estratificación tangencial. Diseño y composición: M. Laura Iribas. Correo: [email protected] Se terminó de imprimir en el mes de septiembre de 2007 en los talleres del Instituto Salesiano de Artes Gráficas, Don Bosco 4053, Buenos Aires, Argentina. ISSN 0328-2767. Reservados todos los derechos. BIOESTRATIGRAFÍA DEL PALEOZOICO SUPERIOR DE AMÉRICA DEL SUR: PRIMERA ETAPA DE TRABAJO HACIA UNA NUEVA PROPUESTA CRONOESTRATIGRÁFICA Convocada por: Paleontólogos de la UBA y de la Fundación Lillo. Auspiciada por: Comité Organizador de la XI Reunião de PaleobotânicosPaleobotânicos y por el Comité Organizador del XIII Simposio Argentino de Paleobotánica y Palinología (Bahía Blanca) Argentina 2006. Comité organizador local Dr. Carlos L. Azcuy CONTENIDO Prólogo .......................................................................................................................................................................................... 5 Introducción ............................................................................................................................................................................. -

Pb-651 PC Srivastava1
The Palaeobotanist 57(2008) : 119-139 0031-0174/2008 $2.00 Some aspects of Palaeozoic pteridophytes of India: A critical reappraisal PRADEEP CHANDRA SRIVASTAVA Department of Botany, Ewing Christian College, University of Allahabad, Allahabad 211 003, India. Email: [email protected] (Received 03 August, 2006; revised version accepted 07 December, 2007) ABSTRACT Srivastava PC 2008. Some aspects of Palaeozoic pteridophytes of India: A critical reappraisal. The Palaeobotanist 57(1-2) : 119-139. The present paper deals with spatial and temporal distribution of pteridophytic megafossils within Palaeozoic plant bearing horizons of India. An attempt has been made to describe the pteridophytic assemblages in various strata of different ages, dynamism of the relative dominance pattern of various types of pteridophytes within each assemblage vis- à-vis the environment they lived in. Critical discussion has been done regarding increasing diversity of Palaeozoic pteridophytic taxa, taxonomic riddles, plant architecture, ecological analysis and phylogenetic implications. Special attention has been given to the floristic regionalism of Lower Carboniferous flora of India in reference to the world palaeophytogeography and occurrence of admixture of Cathaysian, Euramerian and Gondwana pteridophytic elements in palaeoecotone zone near the northwestern boundary of Indian Gondwana Plate. Key-words—Lycopods, Sphenopsids, Pteridophylls, Extra-Peninsular and Peninsular, Pre-Gondwana, Lower Gondwana. Hkkjr ds iqjkthoh VsfjMksQk;Vksa ds dqN igyw -

Article in Press
Türkiye Jeoloji Bülteni Geological Bulletin of Turkey 64 (2021) 267-276 doi: 10.25288/tjb.854704 The First Report of Gangamopteris rajaensis from Rajmahal Gondwana Basin (Jharkhand, India) Arun Joshi1 , Raj Kumar Priya2* 1 Council of Scientific and Industrial Research (CSIR) and SGRR (P.G.) College, Dehradun, India 2 Department of Geology, Sikkim University, Gangtok, Sikkim, India • Geliş/Received: 05.01.2021 • Düzeltilmiş Metin Geliş/Revised Manuscript Received: 13.05.2021 • Kabul/Accepted: 14.05.2021 • Çevrimiçi Yayın/Available online: 29.06.2021 • Baskı/Printed: 25.08.2021 Research Article/Araştırma Makalesi Türkiye Jeol. Bül. / Geol. Bull. Turkey Abstract: The present study deals with the systematic description of macro and miofloral analysis ofGangamopteris rajaensis and Glossopteris indica from the carbonaceous shale-coal bearing sequences of the Rajmahal Open Cast Mine, Rajmahal Basin, Jharkhand, India. The floral diversity, age correlation, and the paleoenvironment of the Barakar Formation were well described. Morphological analysis revealed the reticulate venation pattern, anastomosing of veins, and the absence of the midrib in Gangamopteris rajaensis. The recovered megafloral assemblages of Gangamopteris rajaensis and Glossopteris indica suggest a late early Permian (Artiskian-Kungurian) age for the Barakar strata of Rajmahal coal mine and the prevalence of a moderately warm climate during their deposition. Earlier the species was reported from the Barakar Formation of Damodar and Mahanadi Gondwana basins in India. However, this is the first detailed systematic investigation of this species from the Rajmahal Gondwana Basin, Jharkhand, India. Keywords: Barakar Formation, Early Permian, Gangamopteris rajaensis, Rajmahal Gondwana Basin. INTRODUCTION the Permo-Carboniferous period (Mukhopadhyay The warm and humid climate after the Late et al., 2010). -

Bioestratigrafía Del Pérmico De La Sierra De Los Llanos, La Rioja (Cuenca Paganzo), Sobre La Base De La Megaflora Y Su Correlación Con Áreas Relacionadas
Bioestratigrafía del Pérmico de la Sierra de Los Llanos, La Rioja (Cuenca Paganzo), sobre la base de la megaflora y su correlación con áreas relacionadas Eliana Paula Coturel Tesis para optar por el título de Doctor en Ciencias Naturales Director : Dr. Pedro Raúl Gutiérrez Codirector : Dr. Eduardo Morel Facultad de Ciencias Naturales y Museo Universidad Nacional de La Plata - 2013 - ELIANA PAULA COTUREL – BIOESTRATIGRAFÍA DEL PÉRMICO DE LA SIERRA DE LOS LLANOS … Quiero dedicar esta Tesis a mi mamá, Mónica Edith Emmerich. Creo que no hace falta dar razones de por qué una le dedica su Tesis a su Madre. Tampoco hay, en el rico idioma castellano, palabras suficientes para hacerlo. Agradezco a todos los que hicieron posible el desarrollo de esta Tesis. A mis directores, los Dres. Pedro R. Gutiérrez y Eduardo M. Morel, por la confianza depositada y por permitirme trabajar y aprender bajo su ala. A mis compañeras de equipo, Lucía Balarino y Bárbara Cariglino, por la ayuda y el apoyo que me dieron en todos estos años (nota aquí: Bárbara escaneó las láminas de Glossopteris que usé en el capítulo de morfografía, es importante mencionarlo). Las quiero mucho. Y a Gustavo Correa, por el apoyo en el viaje de campo y el tiempo que compartimos en el Museo. Lo normal sería ver aquí un listado de gente y miles de agradecimientos personales, pero a mí no me parece que sea el lugar (principalmente porque seguro que me olvido de alguien y queda muy feo al estar escrito!!). A cada persona que quiero, que aprecio, se lo hago saber (quizás no con palabras, pero sí con los gestos). -
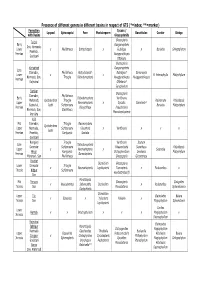
Presence of Different Genera in Different Basins in Respect of GTS
Presence of different genera in different basins in respect of GTS (*=index; **=marker) Formation Cycads / Lyopod Sphenopsid Fern Pteridosperm Benettiales Conifer Ginkgo with Basins Glossopterids Glossopteris Talchir Early Gangamopteris Son, Narmada, Lower x Phyllotheca Botrychiopsis x Rubidgea x Buriadia Ginkgophyton Pranhita, Permian Noeggerothiopis Godabari Ottokaria Glossopteris Karharbari Gangamopteris Late Damodar, Phyllotheca Botrychiopsis* Rubidgea* Samaropsis Lower x x B. heterophylla Platyphyllum Narmada, Son, Trizygia Dichotomopteris Noeggerothiopis Noeggarothiopsis Permian Rajmahal Ottokaria* Euryphyllum Barakar Glossopteris Damodar, Phyllotheca Early Dichotomopteris Vertibraria Mahanadi, Cyclodendron Trizygia Walkomella Rhipidopsis Upper Neomariopteris x Cycads: Barakaria* Rajmahal, lesbii Schizoneura Buriadia Platyphyllum Permian Dizagotheca Pseudocteris Narmada, Son, Stellotheca Macrotaeniopteris Pranhita Kulti Mid Damodar, Trizygia Neomaripteris Cyclodendron Upper Narmada, Schizoneura Gleopteris x Vertibraria x x x lesbii Permian Pranhita, Raniganjia Santalia Godabari Raniganj Trizygia Vertibraria Scutum Late Dichotomopteris Damodar Schizoneura Palaeovittaria Senotheca Rhipidopsis Upper x Neomariopteris x Searsolia Hingir Raniganjia Dictyopteridium Denkania Platyphyllum Permain Damodopteris Mahanadi, Son Phyllotheca Glossopteris Glossotheca Panchet Dicroidium Glossopteris Lower Damodar Trizygia x Neomariopteris Lepidopteris Taenopteris x Podozanites x Triassic Nidpur Schizoneura Kendostrobus(f) Son Marattiopsis Mid Parsora -

Plant Evolution
Conquering the land The rise of plants Ordovician Spores Algae (algal mats) Green freshwater algae Bacteria Fungae Bryophytes Moses? Liverworts? Little body fossil evidence Silurian Wenlock Stage 423-428mya Psilophytes Rhyniopsidsa important later in early Devonian Cooksonia Rhynia Branching stems, flattened sporangia at tips No leaves, no roots short 30 cms rhizoids Zosterophylls Early stem group of Lycopodiophytes Ancestors of Class Lycopsida (clubmosses) Prevalent in Devonian Spores at tips and on branches Lycopsids (?) Baragwanathia with microphylls in Australia Zosterophylls Silurian Cooksonia Development of Soil Fungae Bacteria Algae Organic matter Arthropods and annelids Change in erosion Change in CO2 Devonian Devonian Early Devonian simple structure Rhynie Chert (Rhyniophytes) Trimerophytes First with main shoot Give rise to Ferns and Progymnosperms Up to 3m tall Animal life (mainly arthropods) Late Devonian Forests First true wood (lignin) Forest structure develops (stories) Sphenopsids (Calamites) Lycopsids (Lepidodendron) Seed Ferns (Pteridosperm) Progymnosperm Archaeopteris Cladoxylopsid First vertebrates present Upper Devonian Lycopsida 374-360 mya Leaves and roots differentiated Most ancient with living relatives Megaphylls branching in on plane Photosynthetic webbing Shrub size vertical growth limited (weak) Lateral (secondary) growth (woody) Development of roots Homosporous Heterosporous Upper Devonian Calamites (Sphenopsid) Horestail Sphenophyta (Calamites-Annularia) Devonian Archaeopteris Ur. Devonian - Lr. Carboniferous -

Morphology and Affinities of Glossopteris
MORPHOLOGY AND AFFINITIES OF GLOSSOPTERIS K. R. SURANGE & SHALLA CHANDRA Birbal Sahni Institute of Palaeobotany, Lucknow-226007, India ABSTRACT Reproductive organs of glossopterids, viz., Eretmonia, Glossotlleea, Kendos• trobus, Lidgettonia, Partlla, Russangea, M ooia, Rigbya, Denkania, Venustostrobus, Plumsteadiostrobus, Dietyopteridium and Jambadostrobus are briefly described. Their morphology and affinities are discussed. In the Permian of the southern hemis• phere atleast two distinct orders of gymnosperms, viz., Pteridospermales and Glos• sopteridales were dominating the landscape. genera bore sporangia terminally on ultimate GLOSSOPTERISAustralia were firstleavesrecordedfrom Indiaby Brong•and branches whereas the third genus bore niart in 1828. From 1845 to 1905, sporangia crowded on a cylindrical axis. a number of new species of Glossopteris were discovered from different continents of Eretmonia Du Toit Gondwanaland. From 1905 to 1950 there PI. 1. figs. 3,4; PI. 2, fig. 2; PI. 3, figs. 2,3 were only a few records, but from 1950 onwards Glossopteris again attracted the Fertile scale leaves of Eretmonia (see attention of many palaeobotanists. At first, Chandra & Surange, 1977, text-fig. 1) are of Glos90pteris were regarded as ferns but different shapes and sizes and five species later they turned out to be seed .plants. have been recognized on this basis. The In recent years our knowledge of the fertile bracts are ovate (E. ovoides Surange reproductive structures of glossopterids has & Chandra, 1974), spathulate with acute increased to such an extent that one can apex (E. hingridaensis Surange & Mahesh• get some idea as to what type of plants wari, 1970), triangular (E. emarginata they were. Chandra & Surange, 1977), diamond-shaped (E. utkalensis Surange & Maheshwari, 1970) HABIT and orbicular (E. -

New Glossopterid Polysperms from the Permian La Golondrina Formation (Santa Cruz Province, Argentina): Potential Affinities and Biostratigraphic Implications
Rev. bras. paleontol. 18(3):379-390, Setembro/Dezembro 2015 © 2015 by the Sociedade Brasileira de Paleontologia doi: 10.4072/rbp.2015.3.04 NEW GLOSSOPTERID POLYSPERMS FROM THE PERMIAN LA GOLONDRINA FORMATION (SANTA CRUZ PROVINCE, ARGENTINA): POTENTIAL AFFINITIES AND BIOSTRATIGRAPHIC IMPLICATIONS BÁRBARA CARIGLINO Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, CONICET, Av. Ángel Gallardo 470, C1405DJR, Buenos Aires, Argentina. [email protected] ABSTRACT – Impression fossils of ovuliferous fructifi cations from the Permian La Golondrina Formation in Santa Cruz, Argentina, are described, their affi nities compared, and fi nally, assigned to the Arberiaceae (Glossopteridales). Based on morphological differences from the genera in Arberiaceae, a new taxon is established for some specimens, whereas others are allocated to Arberia madagascariensis (Appert) Anderson & Anderson. This is the fi rst record of Arberiaceae from the La Golondrina Basin. The biostratigraphic implications of the occurrence of this family in this unit are discussed, and suggest that more evidence other than that provided by the megafl oral elements is needed to resolve the age of the constituent members of the La Golondrina Formation. Key words: Arberiaceae, Argentina, biostratigraphy, fructifi cations, Glossopteridales, Gondwana. RESUMO – São descritos impressões fósseis de frutifi cações femininas provenientes da Formação La Golondrina, Permiano de Santa Cruz (Argentina), atribuídas a Arberiaceae (Glossopteridales), segundo a comparação de suas afi nidades. -

1 Relationships of Angiosperms To
Relationships of Angiosperms to 1 Other Seed Plants Seed plants are of fundamental importance both evolution- all gymnosperms (living and extinct) together are not arily and ecologically. They dominate terrestrial landscapes, monophyletic. Importantly, several fossil lineages, Cayto- and the seed has played a central role in agriculture and hu- niales, Bennettitales, Pentoxylales, and Glossopteridales man history. There are fi ve extant lineages of seed plants: (glossopterids), have been proposed as putative close rela- angiosperms, cycads, conifers, gnetophytes, and Ginkgo. tives of the angiosperms based on phylogenetic analyses These fi ve groups have usually been treated as distinct (e.g., Crane 1985; Rothwell and Serbet 1994; reviewed in phyla — Magnoliophyta (or Anthophyta), Cycadophyta, Doyle 2006, 2008, 2012; Friis et al. 2011). These fossil lin- Co ni fe ro phyta, Gnetophyta, and Ginkgophyta, respec- eages, sometimes referred to as the para-angiophytes, will tively. Cantino et al. (2007) used the following “rank- free” therefore be covered in more detail later in this chapter. An- names (see Chapter 12): Angiospermae, Cycadophyta, other fossil lineage, the corystosperms, has been proposed Coniferae, Gnetophyta, and Ginkgo. Of these, the angio- as a possible angiosperm ancestor as part of the “mostly sperms are by far the most diverse, with ~14,000 genera male hypothesis” (Frohlich and Parker 2000), but as re- and perhaps as many as 350,000 (The Plant List 2010) to viewed here, corystosperms usually do not appear as close 400,000 (Govaerts 2001) species. The conifers, with ap- angiosperm relatives in phylogenetic trees. proximately 70 genera and nearly 600 species, are the sec- The seed plants represent an ancient radiation, with ond largest group of living seed plants.