Shoot Density Affects 'Riesling' Grapevines I. Vine Performance
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Alden Lane Nursery Grape Collection
The Alden Lane Nursery Grape Collection Autumn Royal Seedless Large, seedless dark purple to black grape. Vigorous vine that responds best to spur pruning. Appeal rests on large berry size and late maturity. Ripens: Late September to Mid-October Black Monukka Seedless A medium size black seedless grape with a crisp sweet flavor. This is a hardy European variety which can take temperatures as low as 10 °F. A good fresh grape or suitable for raisins. Ripens: August into September Cane or Spur pruning Cabernet Sauvignon Small, round, purple-black grape with many seeds that make a distinctive world-renown red wine. Excellent producer in our area. Ripens late September to October Champagne (Black Corinth or Zante Currants) Although not a true currant, when dried they are referred to as Zante currants and used extensively in baking. It is an old Greek variety. Grape is dark red to black, firm skinned juicy and crunchy with a sweet spicy flavor when ripe. It is seedless. Ripens mid August. Cane or Spur prune Chardonnay For white burgundy style wines. Small round berry. Vigorous vine. Very cold hardy. Ripens: Late August to September Cane pruning Concord Seedless A medium size black seedless version of the classic Concord grape with all its hardiness, vigor, disease resistance and flavor. Excellent for table, juice and jams. Ripens: August Cane or Spur pruning Crimson Seedless Large red seedless table grape. Medium sized clusters of firm, crisp fruit with a sweet flavor. Holds well on vine after ripening. Ripens: October Cane pruning Delight Large. Dark greenish yellow, resembling Thompson Seedless in color. -

New York Concord Grapes Guide
NEW YORK GROWN FOOD GUIDE CONCORD grapes F NYS FARM TO INSTITUTION NEW YORK STATE PMS P 30-7 C CMYK 0, 60, 87, 0 RGB 244, 129, 56 PMS P 30-16 C CMYK 0, 69, 100, 14 RGB 213, 100, 28 PMS P 141-7 C CMYK 87, 0, 81, 42 RGB 0, 116, 69 Western New York’s Concord grape industry exemplifies the very best of this state’s diverse agriculture. By expanding research and marketing opportunities for both Concord grape growers and processors, we can provide a boost to the agricultural community, grow business, and create new jobs in this region and across New York. ANDREW M. CUOMO Governor, New York State As a lifelong visitor to neighboring Chautauqua County, I know there is tremendous local pride in the fact that it is the world’s largest contiguous concord grape growing region. Concord grapes grown across the Lake Erie shoreline are an important part of the diverse agricultural landscape of New York State. KATHY HOCHUL Lieutenant Governor, New York State The juice of the grape is the liquid quintessence of concentrated sunbeams. THOMAS LOVE PEACOCK English Novelist and Poet ii NEW YORK GROWN FOOD GUIDE: CONCORD GRAPES Introduction Farm to Institution New York State (FINYS), Bite into a peanut butter and jelly sandwich or take a swig of pronounced “finesse,” is grape juice and you will instantly recognize the familiar flavor of a collaborative initiative Concord grapes. As the second largest grower of Concord grapes in the nation, New York has just the right climate and soil conditions led by American to grow this popular grape varietal. -

Lake Erie Concord Grape Heritage Area Management Plan
LLakea k e ErieE r i e COONCORDNCORD GRRAPEAPE BEELTLT HHeritageeritage AArearea MManagementanagement PPlanlan peter j. smith & company, inc. “This document was prepared for the New York State Department of State Division of Coastal Resources with funds provided under Title 11 of the Environmental Protection Fund. “ AAcknowledgementscknowledgements The Lake Erie Concord Grape Belt Heritage Area Management Plan was completed with the guidance of various residents and stakeholders interested in the future development and preservation of the Heritage Area. A special thanks to the following participants: Concord Grape Belt Heritage Area Association Town of Westfield and Participating Communities Chautauqua County New York State Office of Parks, Recreation and Historic Preservation New York State Department of State Helen Baran, Concord Grape Belt Heritage Area Association Martha Bills, Town of Westfield David Carr, Village of Westfield, NY Ken Cross, Town of Hanover, NY Patricia Lynch Christina, Town of Pomfret, NY Rick Dunst, Cornell Lake Erie Research and Extension Laboratory Andrew Dufresne, Concord Grape Belt Heritage Area Association Bob Green, Pennsylvania Representative Valeria Ivan, NYS Department of State Marcia Kees, NYS Office of Parks, Recreation and Historic Preservation Kurt Lindstrom, Village of Silver Creek, NY Beth Margolis, Liberty Vineyard and Winery Don McCord, Chautauqua County Dept. of Planning & Economic Development Andrew Nixon, Chautauqua County Visitor’s Bureau Bob Patterson, Town of Portland, NY John Rawlinson, -

Preserving Grapes
Reviewed March 2010 Grapes Christine Jensen, Family and Consumer Science Agent Charlotte Brennand, Food Preservation Specialist Did you know? ! Botanically speaking, grapes are a form of berry. ! European grape varieties include Tokay, Ribier, and Thompson seedless. American grape varieties include Concord, Delaware, Niagara and Isabella. The American grapes tend to be larger and have skins that are easier to slip off, but the seeds are held firmly in the pulp. ! Commercial grape jelly in America is made from Concord grapes. ! Thompson seedless is the most common Concord grapesR grape used to make raisins. SELECTION OF GRAPES High quality grapes are plump, well formed and firmly attached to green, pliable stems. Fully ripe grapes are soft and tender. Grapes showing signs of decay, shriveling, stickiness or dry brittle stems should be avoided. Moldy and wet grapes indicate decay. Unlike some fruits, grapes will not improve or ripen after they have been harvested. They must be at peak quality and sweetness when you purchase or pick them. Green grapes are the sweetest and best flavored when they're yellow-green in color; red varieties when the grapes are predominately red; and the blue-black varieties when the berries have a full rich color. Fresh Facts ! Just before use, wash grape clusters under a gentle spray of water, drain and pat dry. ! Seedless grapes are used whole. For seeded grapes, remove seeds by cutting grapes into halves lengthwise and scooping out seeds with the point of a knife. ! Grapes are easier to peel when they’re frozen. Just rinse frozen grapes in lukewarm water until skins split. -
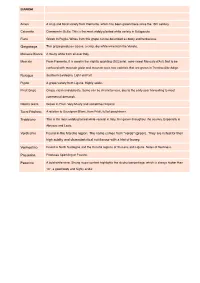
Grape Varieties
BIANCHI Arneis A crisp and floral variety from Piemonte, which has been grown there since the 15th century. Catarratto Common in Sicilia. This is the most widely planted white variety in Salaparuta. Fiano Grown In Puglia. Wines from this grape can be described as dewy and herbacious. Garganega This grape produces Soave, a crisp, dry white wine from the Veneto. Malvasia Bianca A musty white from all over Italy. Moscato From Piemonte, it is used in the slightly sparkling (frizzante), semi-sweet Moscato d'Asti. Not to be confused with moscato giallo and moscato rosa, two varietals that are grown in Trentino Alto-Adige. Nuragus Southern Sardegna. Light and tart. Pigato A grape variety from Liguria. Highly acidic. Pinot Grigio Crispy, clean and delicate. Some can be character-less, due to the early year harvesting to meet commercial demands. Ribolla Gialla Grown in Friuli. Very Musty and sometimes tropical. Tocai Friulano A relation to Sauvignon Blanc, from Friuli, full of peachiness. Trebbiano This is the most widely planted white varietal in Italy. It is grown throughout the country, Especially in Abruzzo and Lazio. Verdicchio Found in the Marche region. The name comes from "verde" (green). They are noted for their high acidity and characteristical nut flavour with a hint of honey. Vermentino Found in North Sardegna and the Coastal regions of Toscana and Liguria. Notes of freshness. Passerina Produces Sparkling or Passito. Pecorino A bold white wine. Strong sugar content highlights the alcohol percentage, which is always higher than 13°, a good body and highly acidic. ROSSI Aglianico Considered the "noble varietal of the south," grown in Campania and Basilicata. -

Medicines and Inedibles : Wine and Grape Juice
SPECIAL PESACH EDITION ww ww VOLUME u y / NO. 6 ADAR 5767 / MARCH 2007 s xc THEDaf HaKashrus A MONTHLY NEWSLETTER FOR THEU RABBINIC FIELD REPRESENTATIVE UP LABEL NAME BRAND NAME : WINE AND GRAPE JUICE - Aloxe Corton (Not Mevushal) Herzog French Asti (Mevushal) Bartenura MEVUSHAL OR NON-MEVUSHAL? Asti (Mevushal) Rashi Barbera D’Alba (Mevushal) Rashi Select DAF NOTES: Barbera D’Asti (Mevushal) Bartenura The OU maintains the position of the Igros Moshe that heating Barolo (Mevushal) Rashi Select grape juice or wine to 175° F renders the product Mevushal and Baron Rothschild (Not Mevushal) Herzog French thereby permissible to be handled by non-Jews. Beaujolais Nouveau (Mevushal) Herzog French All wine products bearing the OU symbol indicating Mevushal Beaujolais Villages (Mevushal) Herzog French have been heated to a minimum 175° F. Beaujolais Villages Blanc (Not Mevushal) Herzog French All OU grape juice is pasteurized at a minimum 175° F. The only Black Muscat (Mevushal) Baron Herzog exception to this rule is the Kedem 1.5 liter glass grape juice whose Wine label clearly states “not Mevushal.” This product is pasteurized at Blanc De Blancs Brut (Mevushal) Herzog French lower temperatures. Blanco (Not Mevushal) Ramon Cardova Blush Chablis (1.5 L) (Not Mevushal) Kedem Estates There are other opinions requiring higher temperatures. The OU Blush Chablis (750 Ml) (Mevushal) Kedem Estates does not follow these. Blush Concord (1.5L) (Not Mevushal) Kedem The position of the OU Poskim is that, although there is a Hidur Blush Concord (750 Ml) (Mevushal) Kedem to use non-mevushal wine for Mitzva purposes (Kiddush, Seder Blush Concord (Mevushal) Tam Pree Light Cups), mevushal wine may still be used lechatchila. -

History of Grape Growing in the Eastern United States
History of Grape Growing in the Eastern United States By Fred Elmer Gladwin 1931 Reproduced in 2007 from the original work which was published in eleven parts in Rural New Yorker in 193 1. Fred Elmer Gladwin (1 877-1940) was a well regarded expert in viticulture. In 1909 he was appointed a special agent of the New York State Agricultural Experiment Station (NYSAES) and put in charge of the Vineyard Laboratory at Fredonia, NY which came into existence that same year. Later he was made associate in research at the NYSAES, and in 1936 was made chief in research in Pomology, a position he held at the time of his death. His entire period of service with the Experiment Station was spent at the Vineyard Laboratory. This series of columns he authored in 1931 on the history of grape growing in the Eastern United States has been brought together here in an effort to give more prominence to a small part of his work. The citations for the work as it originally appeared are listed here: Gladwin, F.E., "A History of Grape Growing in Eastern United States: Part I," Rural New Yorker, vol.XC, no.5 156,18 April 1931, page 485-486. Gladwin, F.E., "A History of American Grape-growing: Part 11," Rural New Yorker, vol.XC, no.5 157,25 April 1931, page 5 13-5 14. Gladwin, F.E., "A History of Grape-growing in Eastern United States: Part 111," Rural New Yorker, vol.XC, no.5158,2 May 1931, page 543,545. Gladwin, F.E., "A History of Grape-growing in the Eastern United States: Part IV," Rural New Yorker, vol.XC, no.5 159,9 May 1931, page 565. -

Potassium in Viticulture and Enology Russell Moss (Graduate Research Assistant, Virginia Tech) May 2016
Potassium in viticulture and enology Russell Moss (Graduate Research Assistant, Virginia Tech) May 2016 Introduction Potassium (K+) is of interest to Virginia vintners, as many of the soils upon which vineyards have been planted have high exchangeable K+ levels and while excessive K+ does not cause “toxicity”, high soil concentrations can interfere with uptake of certain other nutrients. Furthermore, high K+ concentrations in grapes has potentially negative implications for wine acid profiles, pH and color stability. In a warm climate, such as Virginia, titratable acidity is known to degrade rapidly from véraison until harvest. This leads to wines which can be out of balance and considered “flabby” (i.e. lacking acidity), and such wines may also have significantly elevated pH. Tartaric acid can be added to the must to reduce pH; however, high concentrations of K+ in the juice can reduce the effectiveness of the acid addition. Therefore, it is to the advantage of the vintner to minimize the negative effects of K+ in the juice. This review will discuss: 1. The forms in which K+ is found in the soil 2. Mode of uptake and transport of K+ in the plant 3. Function of K+ in the plant 4. Environmental factors effecting uptake and transport of K+ 5. Viticultural practices which influence K+ uptake and transport 6. Enological consequences of high K+ in the juice K+ in the soil Potassium is generally the most abundant cation found in mineral soils (Reitemeier 1951). However, much of the K+ found in soils isn’t easily exchangeable and is held in primary minerals which only release the K+ gradually over considerable periods of time (Brady and Weil 1996). -

Orape, Raisin, and Wine Production in the United States
ORAPE, RAISIN, AND WINE PRODUCTION IN THE UNITED STATES. By GEORGE C. HUSMANN, Expert in Charge of Viticultural Investigations y Bureau of Plant Industry. THE GRAPE-GROWING INDUSTRY. EARLY HISTORY. When America was discovered the wild vine was so prominent a feature of the vegetation that the name Yineland was more than once applied to the country. Considerable wine was produced from a native grape in Florida as early as 1564. The London Company planted vineyards in Virginia prior to 1620, and many succeeding attempts at grape growing were made by William Penn and by Ger- man and Swiss settlers. Of more recent attempts to cultivate the vine on the Atlantic coast, the first were confined to European varieties, and were not successful. The Mission Fathers in California were the first to successfully grow the European grape in the United States. They grew grapes at the missions for their own use only, the work being principally done by Indians. They had but one variety, v/hich is still largelj^ grown, and is known by the name of Mission. It is first heard of as introduced into Mexico in 1520. Chronologically, it was brought to the California missions, as follows: San Diego, 1769; San Gabriel, 1771; Los Angeles, 1781; and Santa Barbara, 1786. The Mission vine planted at Montecito, Cal., in 1795, was exhibited at the Centennial Exposition in Philadelphia. It was 18 inches in diameter, and in one season had produced over 5 tons of grapes. From the missions, the viticultural pioneers received their inspira- tion as well as their start of cuttings NEW ERA IN GRAPE GROWING. -

WINERY OVERVIEW a Name Steeped in Tradition, Manischewitz Is A
WINERY OVERVIEW All Manischewitz wines are certified kosher. Manischewitz A name steeped in tradition, Manischewitz is a traditional also produces some varieties that are kosher for Passover. favorite in the Kosher wine category. From the crushing of Passover Requirements: the grapes to the final bottling, the Manischewitz • Kosher for Passover wines are produced using real commitment is reflected in every step of the Manischewitz cane sugar, instead of corn syrup. wine production. Along the way, the Union of Orthodox • Kosher for Passover items are produced four months Jewish Congregations of America supervises every aspect prior to the holiday. to certify that Manischewitz wines are 100% kosher, and that respected designation honors each and every bottle. • All wines, except Elderberry and Cream Peach are offered as kosher for Passover items. KEY SELLING POINTS See the chart for a listing of varietals, sizes, and kosher 1. The #1 kosher wine brand in the U.S.1 certification. (See back for details) 2. Available in kosher for Passover (during the Passover holiday selling season). WINEMAKER INFORMATION Joyce Magin began her career in the wine industry as a lab 3. Consumers say that the strong heritage and tradition technician at Widmer Wine Cellars, working her way up to of Manischewitz make it an integral part of Jewish the position of winemaker over the course of more than 20 holiday celebrations!2 years with the company. Upon the consolidation of East Coast 1IRI Multi Outlet + Convenience, L52 wks ending 2/23/14 production, she joined the team of winemakers in Canandaigua, 2Insights in Marketing Qualitative Consumer Research, September 2006 and now oversees the production of Manischewitz and bulk contract wines, totaling over 4,000,000 gallons. -

The Science of Wine: Washington State University Scientists and the Development of the Washington Wine Industry, 1937-1992
THE SCIENCE OF WINE: WASHINGTON STATE UNIVERSITY SCIENTISTS AND THE DEVELOPMENT OF THE WASHINGTON WINE INDUSTRY, 1937-1992 By Cynthia Stewart Kaag A dissertation submitted in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY WASHINGTON STATE UNIVERSITY Department of History December 2008 To the Faculty of Washington State University: The members of the Committee appointed to examine the dissertation of CYNTHIA STEWART KAAG find it satisfactory and recommend that it be accepted. _____________________________________ (Chair) _____________________________________ _____________________________________ ii TABLE OF CONTENTS ACKNOWLEDGMENTS iv ABSTRACT v ABBREVIATIONS vi GLOSSARY vii LIST OF FIGURES x DEDICATION xi INTRODUCTION 12 CHAPTER 1: GRAPES AND WINES IN WASHINGTON FROM 1937 TO 1959 27 CHAPTER 2: A GROWING INDUSTRY: THE 1960S 41 CHAPTER 3: CORRESPONDENCE: A CAREER IN A YEAR 74 CHAPTER 4: GRANTSMANSHIP IN THE 1970S 100 CHAPTER 5: FROM PROJECT TO PUBLICATION 150 CHAPTER 6: MATURING WINES AND WINES: THE 1980S 164 CHAPTER 7: COMING OF AGE: THE EARLY1990S AND AFTER 182 BIBLIOGRAPHY 192 APPENDICES 201 Appendix A: WSU Theses Related to Wine, 1948-1992 Appendix B: WSU Wine Publications Appendix C: Biographical Sketches Appendix D: Grape Varieties Recommended for Washington State iii ACKNOWLEDGMENTS For their support and encouragement I am deeply indebted to the members of my committee, Drs. Orlan Svingen, Carolyn Ross, and most especially my advisor Jerry Gough. I was especially privileged to have the opportunity to take classes and seminars with Dr. Gough, during which he helped me toward an understanding of the role and significance of science and technology in our history and culture. I also benefited very much from classes with Drs. -

'He Sowed; Others Reaped': Ephraim Wales Bull and The
"He Sowed; Others Reaped": Ephraim Wales Bull and the Origins of the ‘Concord’ Grape Edmund A. Schofield While Emerson and his colleagues were designing a philosophy for the unique needs of an expanding nation, one of their townsmen was quietly developing a grape to match the demands of its rigorous physical environment The Origins and Spread of "The Vine" era, grapes and wine were of considerable In their peregrinations over the ages, the importance to Middle Eastern and Mediterra- grape and mankind have crossed paths, have nean peoples. Thousands of years ago the even trod the same path, many times. East Egyptians were well acquaintcd with the use and West, for millennia, they have followed and properties of wine, which their traditions similar routes of history, myth, and ro- say were revealed to them by Osiris. Their mance-first in the Northern Hemisphere, in chief vineyards were planted on the banks of Asia, in Europe, in North America, and then, the Nile. Joseph’s dream, described in Gene- within the last few centuries, in the Southern sis, gives evidence that the vine was culti- Hemisphere as well. Companion to mankind vated in Egypt at least eighteen hundred years from dimmest antiquity, the grape has been before Christ. Grape seeds have been found one of mankind’s most important, most es- with mummies in Egyptian tombs that are at teemed fruits. least three thousand years old, and details of In the West, the story of the grape has been grape growing appear in mosaics of the Fourth largely the story of Vitis vinifera Linnxus- Dynasty of Egypt (2440 B.C.) and later.