Spatial and Temporal Variation of Anchovy Predationby Albacore And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

DYNAMIC HABITAT USE of ALBACORE and THEIR PRIMARY PREY SPECIES in the CALIFORNIA CURRENT SYSTEM Calcofi Rep., Vol
MUHLING ET AL.: DYNAMIC HABITAT USE OF ALBACORE AND THEIR PRIMARY PREY SPECIES IN THE CALIFORNIA CURRENT SYSTEM CalCOFI Rep., Vol. 60, 2019 DYNAMIC HABITAT USE OF ALBACORE AND THEIR PRIMARY PREY SPECIES IN THE CALIFORNIA CURRENT SYSTEM BARBARA MUHLING, STEPHANIE BRODIE, MICHAEL JACOX OWYN SNODGRASS, DESIREE TOMMASI NOAA Earth System Research Laboratory University of California, Santa Cruz Boulder, CO Institute for Marine Science Santa Cruz, CA CHRISTOPHER A. EDWARDS ph: (858) 546-7197 Ocean Sciences Department [email protected] University of California, Santa Cruz, CA BARBARA MUHLING, OWYN SNODGRASS, YI XU HEIDI DEWAR, DESIREE TOMMASI, JOHN CHILDERS Department of Fisheries and Oceans NOAA Southwest Fisheries Science Center Delta, British Columbia, Canada San Diego, CA STEPHANIE SNYDER STEPHANIE BRODIE, MICHAEL JACOX Thomas More University, NOAA Southwest Fisheries Science Center Crestview Hills, KY Monterey, CA ABSTRACT peiods, krill, and some cephalopods (Smith et al. 2011). Juvenile north Pacific albacore Thunnus( alalunga) for- Many of these forage species are fished commercially, age in the California Current System (CCS), supporting but also support higher-order predators further up the fisheries between Baja California and British Columbia. food chain, such as other exploited species (e.g., tunas, Within the CCS, their distribution, abundance, and for- billfish) and protected resources (e.g., marine mammals aging behaviors are strongly variable interannually. Here, and seabirds) (Pikitch et al. 2004; Link and Browman we use catch logbook data and trawl survey records to 2014). Effectively managing marine ecosystems to pre- investigate how juvenile albacore in the CCS use their serve these trophic linkages, and improve robustness of oceanographic environment, and how their distributions management strategies to environmental variability, thus overlap with the habitats of four key forage species. -
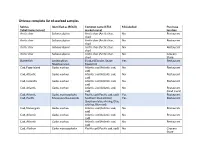
Ottawa: Complete List of Seafood Samples
Ottawa: complete list of seafood samples Sold as Identified as (BOLD) Common name (CFIA Mislabelled Purchase (label/menu/server) market name) location Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Restaurant char) Arctic char Salverus alpirus Arctic char (Arctic char, No Grocery char) Store Butterfish Lepidocybium Escolar (Escolar, Snake Yes Restaurant flavobrunneum Mackerel) Cod, Fogo Island Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) (food truck) Cod, Atlantic Gadus macrocephalus Pacific cod (Pacific cod, cod) Yes Restaurant Cod, Pacific Micromesistius australis Southern blue whiting Yes Restaurant (Southern blue whiting, Blue whiting, Blue cod) Cod, Norwegian Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Atlantic Gadus morhua Atlantic cod (Atlantic cod, No Restaurant cod) Cod, Alaskan Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, Pacific Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, North Atlantic Gadus macrocephalus Pacific cod (Pacific cod, cod) Yes Restaurant Cod Gadus macrocephalus Pacific cod (Pacific cod, cod) No Grocery Store Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Grocery cod) Store Cod, Icelandic Gadus morhua Atlantic cod (Atlantic cod, No Grocery cod) Store Euro Bass Gadus morhua Atlantic cod (Atlantic cod, Yes Restaurant cod) Grouper Epinephelus diacanthus Spinycheek grouper (n/a) Yes – E. -

Predator/Prey Interactions, Competition, Multi
Current understanding of trophic dynamics In the Mid-Atlantic Bluefish diets, Mid Atlantic and Southern New England, NEFSC surveys 100% All others All Flatfish 90% Unid fish Unid Unid fish Unid fish Unid fish Scup 80% Scup Unid fish Scup Bluefish Butterfish Bluefish 70% Mackerel Bluefish Butterfish Butterfish Hakes 60% Loligo Drums Butterfish Unid Squid Ocean pout Round herring 50% Butterfish Ctenophores Menhaden Loligo Loligo Loligo 40% Loligo Unid Squid Round herring Illex Bay anchovy Unid Squid Unid Squid Silver anchovy Round herring Round herring 30% Bay anchovy Menhaden Unid herring Bay anchovy Menhaden Bay anchovy Striped anchovy 20% Silver anchovy Unid anchovies Bay anchovy Striped anchovy Unid anchovies 10% Striped anchovy Unid anchovies Sand lances Unid anchovies Sand lances Sand lances Sand lances 0% 1977-86 1987-96 1997-2006 2007-2013 Food web models partition mortality 100% 90% 80% If Pred > F, 70% Pred Re-evaluate 60% constant M 50% 40% Proportion of total Mortality total Proportion of F 30% 20% 10% Proportion of total mortality total of Proportion 0% Longnose skate P. Halibut W. Pollock Squids Gaichas et al. 2010 Fishing Predation Other Link et al. 2008. The Northeast U.S. continental shelf Energy Modeling and Analysis exercise (EMAX): Ecological network model development and basic ecosystem metrics. Journal of Marine Systems 74: 453-474 Intermediate tactical multispecies models An intermediate-complexity tactical ecosystem assessment tool combines: Standard stock assessment Ecosystem considerations • Structured population -

Invasive Catfish Management Strategy August 2020
Invasive Catfish Management Strategy August 2020 A team from the Virginia Department of Game and Inland Fisheries uses electrofishing to monitor invasive blue catfish in the James River in 2011. (Photo by Matt Rath/Chesapeake Bay Program) I. Introduction This management strategy portrays the outcomes of an interactive workshop (2020 Invasive Catfish Workshop) held by the Invasive Catfish Workgroup at the Virginia Commonwealth University (VCU) Rice Rivers Center in Charles City, Virginia on January 29-30, 2020. The workshop convened a diverse group of stakeholders to share the current scientific understanding and priority issues associated with invasive catfishes in Chesapeake Bay. The perspectives shared and insights gained from the workshop were used to develop practical, synergistic recommendations that will improve management and mitigate impacts of these species across jurisdictions within the watershed. Blue catfish (Ictalurus furcatus) and flathead catfish (Pylodictis olivaris) are native to the Ohio, Missouri, Mississippi, and Rio Grande river basins, and were introduced into the Virginia tributaries of Chesapeake Bay in the 1960s and 1970s to establish a recreational fishery. These non-native species have since spread, inhabiting nearly all major tributaries of the Bay watershed. Rapid range expansion and population growth, particularly of blue catfish, have led to increasing concerns about impacts on the ecology of the Chesapeake Bay ecosystem. 1 Chesapeake Bay Management Strategy Invasive Catfish Blue and flathead catfishes are long-lived species that can negatively impact native species in Chesapeake Bay through predation and resource competition. Blue catfish are generalist feeders that prey on a wide variety of species that are locally abundant, including those of economic importance and conservation concern, such as blue crabs, alosines, Atlantic menhaden, American eels, and bay anchovy. -

Yellowfin Tuna
Ahi yellown tuna (Thunnus albacares) is one of two Islands. species known in Hawaii simply as Fishing Methods: intermediaries on all islands, or di- ahi. Similar in general appearance rectly to wholesalers and retailers, or it may be shared with family and to bigeye tuna (the other species - known as ahi friends. Most ahi is sold fresh, but men. A large part of the commercial surpluses caught during the peak be recognized by its more torpedo catch (44%) is harvested by longline shaped body, smaller head and eyes. summer season are sometimes dried boats, which may search for tuna and smoked. In Hawaii, shibi is another name up to 800 nautical miles from port and set hooks in deep waters. Yel- Quality to depths below 600 ft. Landings by either bigeye or albacore tuna. Al- lengthen with age. the island of Hawaii, can be sub- stantial (36%) in some years. Troll- Seasonality & How ers contribute most of the remain- does not retain the beautiful natu- They Are Caught der (20%) of the commercial catch ral red color as long as bigeye. The - Availability and Seasonality: - Caught year-round in Hawaii’s wa- ing tournaments held in Hawaii. method, care in handling and other Distribution: abundant during the summer sea- The longline catch and some of the son (May-September). There are handline (ika-shibi) catch of ahi is species. Noticeable changes occur auction. The majority of the hand- Hawaii. ocean surface temperatures and line catch is sold to wholesalers and other oceanographic conditions fa- intermediary buyers on the island of surface during the summer season vor the migration of ahi schools to are susceptible to a quality defect The troll catch may be marketed known as “burnt tuna”. -

Albacore Tuna Have fl Uctuated Considerably from Year To
Tuna [211] 86587_p211_220.indd 211 12/30/04 4:53:37 PM highlights ■ The catches of Pacifi c bluefi n tuna and North Pacifi c albacore tuna have fl uctuated considerably from year to Ocean year, but no upward or downward trends are apparent for either species. and ■ Increasing the age at entry of Pacifi c bluefi n into the fi shery might increase the yields per recruit of that Climate species. ■ The status of North Pacifi c albacore is uncertain, but most scientists believe that greater harvests of that species Changes would not be sustainable. [212] 86587_p211_220.indd 212 12/30/04 4:53:38 PM background The Inter-American Tropical Tuna Commission (IATTC) studies the tunas of the eastern Pacifi c Ocean (EPO), defi ned for its purposes as the area bounded by the coastline of North, Central, and South America, 40ºN, 150ºW, and 40ºS. The IATTC staff maintains records for most of the vessels that fi sh at the surface for skipjack tuna (Katsuwonus pelamis), yellowfi n tuna (Thunnus albacares), bigeye tuna (T. obesus), and Pacifi c bluefi n tuna (T. orientalis) in the EPO. Pacifi c bluefi n and albacore tuna (T. alalunga) are the tunas most relevant to the region of interest to PICES. Pacifi c bluefi n tuna Spawning of Pacifi c bluefi n apparently takes place only Age-1 and older fi sh are caught by purse seining, in the western Pacifi c Ocean (WPO). Some juvenile mostly during May-September between about 30°- bluefi n move from the WPO to the EPO, and then later 42°N and 140°-152°E. -

Seafood Guide
eat It’s good for you! What pregnant and breastfeeding women and parents of young children need to know. Fish are nutritious and most are very How can you safely safe to eat. eat fish? • Fish have protein and healthy fats, called omega-3s, which are not • Eat a variety of fish that are lower found in other meats. in mercury. • Omega-3s are good for your heart • Eat the amounts of fish shown on and brain. the other side of this pamphlet. • The nutrients in fish are especially • Eat only the flesh or meat of important as your baby develops the fish. Throw away the bones, during pregnancy, throughout head, guts, fat, and skin. breastfeeding, and as your young • Avoid shark, swordfish, tilefish, or child grows. king mackerel. They are highest in • Some fish may contain a chemical mercury. called mercury. Too much mercury • Avoid raw and undercooked in your diet can be harmful. It’s fish and shellfish. best to eat fish that are lower in mercury. For more information about mercury in your fish, visit the Environmental Protection Agency — Fish Advisory at www.epa.gov/choose-fish-and-shellfish-wisely. choose safe Follow these tips to enjoy the health benefits of eating fish low in mercury and high in omega-3s. 1. Safe to Eat 2. Do Not Eat Eat fish from the list below 2 to 3 These fish are high in mercury. times a week. Choose fish from stores • Shark • King Mackerel or restaurants. • Swordfish • Tilefish • For women, eat about 8 to 12 ounces a week total. -

Mid-Atlantic Forage Species ID Guide
Mid-Atlantic Forage Species Identification Guide Forage Species Identification Guide Basic Morphology Dorsal fin Lateral line Caudal fin This guide provides descriptions and These species are subject to the codes for the forage species that vessels combined 1,700-pound trip limit: Opercle and dealers are required to report under Operculum • Anchovies the Mid-Atlantic Council’s Unmanaged Forage Omnibus Amendment. Find out • Argentines/Smelt Herring more about the amendment at: • Greeneyes Pectoral fin www.mafmc.org/forage. • Halfbeaks Pelvic fin Anal fin Caudal peduncle All federally permitted vessels fishing • Lanternfishes in the Mid-Atlantic Forage Species Dorsal Right (lateral) side Management Unit and dealers are • Round Herring required to report catch and landings of • Scaled Sardine the forage species listed to the right. All species listed in this guide are subject • Atlantic Thread Herring Anterior Posterior to the 1,700-pound trip limit unless • Spanish Sardine stated otherwise. • Pearlsides/Deepsea Hatchetfish • Sand Lances Left (lateral) side Ventral • Silversides • Cusk-eels Using the Guide • Atlantic Saury • Use the images and descriptions to identify species. • Unclassified Mollusks (Unmanaged Squids, Pteropods) • Report catch and sale of these species using the VTR code (red bubble) for • Other Crustaceans/Shellfish logbooks, or the common name (dark (Copepods, Krill, Amphipods) blue bubble) for dealer reports. 2 These species are subject to the combined 1,700-pound trip limit: • Anchovies • Argentines/Smelt Herring • -

Fish for Your Health™ Advice for Pregnant Or Nursing Women, Women That Will Become Pregnant, and Children Under 6 Years of Age
Fish for Your Health™ Advice for pregnant or nursing women, women that will become pregnant, and children under 6 years of age 1. Eat fish – Health experts recommend that women eat 8-12 ounces/week (weight before cooking) of fish. Children, ages 2-6, should eat at least 2 ounces/week. As a reference, 3 ounces of fish is about the size of a deck of cards. Women that eat fish which contains omega-3 fatty acids (EPA & DHA) will pass these nutrients to their babies and support healthy brain and eye development. Best Choices: Eating six ounces/week of the following fish provides the recommended amounts of healthy fats and will minimize your baby’s exposure to pollutants: salmon (wild or farm-raised), rainbow trout (farm-raised), herring, mackerel (Atlantic, Jack, chub), sardine, shad (American), whitefish. 2. Before eating recreationally-caught fish, check our Fish4Health website below for your State’s fish consumption advisory and avoid eating fish that is heavily contaminated with pollutants. If a fish that you caught is not listed in the advisory, then eat no more than 1 meal per month. If you are unsure about the safety of the fish that you caught, be safe - ‘catch-and-release’. 3. Minimize your exposure to pollutants in commercial fish - follow the advice given below. (Ex: If you eat 4 ounces of albacore tuna, then don’t eat any other fish from this category until the following week.) Level of Maximum Mercury Amount for Commercial Fish Species or PCBs** Adults to Eat anchovy, butterfish, catfish (farm-raised), clam, cod, crab (Blue, King -

ANNEX 14 STOCK ASSESSMENT of PACIFIC BLUEFIN TUNA (Thunnus
FINAL ISC/18/ANNEX/14 ANNEX 14 18th Meeting of the International Scientific Committee for Tuna and Tuna-Like Species in the North Pacific Ocean Yeosu, Republic of Korea July 11-16, 2018 STOCK ASSESSMENT OF PACIFIC BLUEFIN TUNA (Thunnus orientalis) IN THE PACIFIC OCEAN IN 2018 REPORT OF THE PACIFIC BLUEFIN TUNA WORKING GROUP July 2018 1 FINAL Left Blank for Printing FINAL Table of Contents 2018 Pacific Bluefin Tuna Stock Assessment ...........................................................................5 ISC PBFWG ...............................................................................................................................5 EXECUTIVE SUMMARY MAY 2018 ....................................................................................5 1.0 INTRODUCTION .............................................................................................................24 2.0 BACKGROUND ON BIOLOGY AND FISHERIES .......................................................25 2.1 Biology ...................................................................................................... 25 2.1.1 Stock Structure .............................................................................................. 25 2.1.2 Reproduction .................................................................................................. 25 2.1.3 Distribution and Movements ........................................................................ 25 2.1.4 Growth .......................................................................................................... -

1 Updated Standardized CPUE of Atlantic Bluefin Tuna Caught
SCRS/08/100 Updated standardized CPUE of Atlantic bluefin tuna caught by the Spanish baitboat fishery in the Bay of Biscay (Eastern Atlantic). Time series from 1975 to 2007. Enrique Rodriguez-Marin1, Mauricio Ortiz2, Cristina Rodríguez-Cabello1 and Santiago Barreiro1 SUMMARY Updated standardized relative abundance indices by age are presented for bluefin tuna baitboat fishery in the Bay of Biscay from 1975 to 2007. Standardization was carried out using generalized linear mixed models. Catch and effort data on bluefin tuna were available from catches by trip; this catch by commercial category was converted to catch at age by applying seasonal age length keys to the length distribution by commercial category. In this update the age length keys of summer of the entire study period were reviewed. Age class was included as a fixed factor within model specifications because the fishery operates on all available stock fractions, and age determination of the tuna landings is done after removal of the catch. Splitting of the catches into age classes introduces a large number of zero values, so data were modeled using the delta- lognormal model. The model finally selected included the following explanatory factors: Year, Age, Month and Year × Age fixed factors, plus a selection of other factors contributing with a significant percentage of the total explained deviance in the aggregated model. All Year interactions besides the Year × Age factor were considered as random. CVs of present standardized index are less variable than the previous standardization time series from 1975 to 2004, but still some variability was found for the last years when the vessels built more recently and larger are included in the analysis. -

C1. Tuna and Tuna-Like Species
163 C1. TUNA AND TUNA-LIKE SPECIES exceptional quality reached US$500 per kg and by Jacek Majkowski * more recently even more, but such prices referring to very few single fish do not reflect the INTRODUCTION situation with the market. Bigeye are also well priced on the sashimi markets. Although The sub-order Scombroidei is usually referred to yellowfin are also very popular on these markets, as tuna and tuna-like species (Klawe, 1977; the prices they bring are much lower. For Collette and Nauen, 1983; Nakamura, 1985). It is canning, albacore fetch the best prices due to composed of tunas (sometimes referred to as true their white meat, followed by yellowfin and tunas), billfishes and other tuna-like species. skipjack for which fishermen are paid much less They include some of the largest and fastest than US$1 per kg. The relatively low prices of fishes in the sea. canning-quality fish are compensated by their The tunas (Thunnini) include the most very large catches, especially in the case of economically important species referred to as skipjack and yellowfin. Longtail tuna principal market tunas because of their global (T. tonggol) is becoming increasingly important economic importance and their intensive for canning and the subject of substantial international trade for canning and sashimi (raw international trade. The consumption of tuna and fish regarded as delicacy in Japan and tuna-like species in forms other than canned increasingly, in several other countries). In fact, products and sashimi is increasing. the anatomy of some tuna species seems to have The tunas other than the principal market species been purpose-designed for canning and loining.