(Terminalia Superba Engl. & Diels) in the Framework Of
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
GUINEE - PREFECTURE DE NZEREKORE Carte De La Population Et Des Structures Sanitaires 10 Mars 2021
GUINEE - PREFECTURE DE NZEREKORE Carte de la population et des structures sanitaires 10 mars 2021 BEYLA KOUANKAN BOOLA Boola Kognea Maghana Lomou Ouinzou Kogbata FOUMBADOU SEREDOU Boma Nord Gbana KOROPARA Lomou Gounangalaye Saouro Kpamou Koni Koropara Lokooua Alaminata Foozou Nyema Nord Kabieta Bassa´ta Koroh Keleta Ouleouon WOMEY Pampara Suobaya Bowe Nord NZEBELA Keorah Womey kerezaghaye Zenemouta Beneouli Selo MACENTA Loula Yomata Koule Komata KOULE Pale Yalakpala Tokpata Goueke Voumou LAINE Tamoe Niaragbaleye Kpoulo Pouro Kolata Maoun Wouyene PALE Zogbeanta GOUECKE Dapore Soulouta SOULOUTA Konipara Gouh Nona Banzou Tamoe Kola Kpagalaye Nyenh Koronta Youa Gbouo Kelemanda Zogotta Mahouor Koloda KOBELA KOYAMAH Kpaya Souhoule Kelouenta Poe KOKOTA Filikoele Koola Yleouena Batoata Gbadiou Niambara LOLA SAMOE Niema Komou Niarakpale Gbamba Banota Kpaya YALENZOU Boma Samoe Moata Kounala LOLA CTRE Koule Sud Boma Sud Horoya BOWE NZEREKORE CTRE Dorota Nyalakpale Bangoueta Gonia Nyen Commercial Toulemou Nzerekore Konia Kotozou Wessoua Aviation Mohomuo YOMOU Gbottoye Yalenzou Secteur zaapa Zao Yalenzou Kerema Gbenemou Lomou Centre centre Toulemou POPULATION PAR SOUS-PREFECTURE NZOO Gbenedapah SOUS-PREFECTURE POPULATION 2021 Foromopa BOSSOU Teyeouon Bounouma 27 605 Bounouma Kankore Gouecke 24 056 centre DISTANCES DES DISTRICTS SANITAIRES DE NZEREKORE Kobela 19 080 Centre de santé Distance N’Zérékoré_CS Poste de santé Distance CS_PS Koropara 23 898 KOULE SUD 9KM NYEMA 10 KM Sehipa Koule 23 756 C S SAMOE 12 KM NYAMPARA PELA 9 KM BOUNOUMA Thuo centre -

Guinea : Reference Map of N’Zérékoré Region (As of 17 Fev 2015)
Guinea : Reference Map of N’Zérékoré Region (as of 17 Fev 2015) Banian SENEGAL Albadariah Mamouroudou MALI Djimissala Kobala Centre GUINEA-BISSAU Mognoumadou Morifindou GUINEA Karala Sangardo Linko Sessè Baladou Hérémakono Tininkoro Sirana De Beyla Manfran Silakoro Samala Soromaya Gbodou Sokowoulendou Kabadou Kankoro Tanantou Kerouane Koffra Bokodou Togobala Centre Gbangbadou Koroukorono Korobikoro Koro Benbèya Centre Gbenkoro SIERRA LEONE Kobikoro Firawa Sassèdou Korokoro Frawanidou Sokourala Vassiadou Waro Samarami Worocia Bakokoro Boukorodou Kamala Fassousso Kissidougou Banankoro Bablaro Bagnala Sananko Sorola Famorodou Fermessadou Pompo Damaro Koumandou Samana Deila Diassodou Mangbala Nerewa LIBERIA Beindou Kalidou Fassianso Vaboudou Binemoridou Faïdou Yaradou Bonin Melikonbo Banama Thièwa DjénédouKivia Feredou Yombiro M'Balia Gonkoroma Kemosso Tombadou Bardou Gberékan Sabouya Tèrèdou Bokoni Bolnin Boninfé Soumanso Beindou Bondodou Sasadou Mama Koussankoro Filadou Gnagbèdou Douala Sincy Faréma Sogboro Kobiramadou Nyadou Tinah Sibiribaro Ouyé Allamadou Fouala Regional Capital Bolodou Béindou Touradala Koïko Daway Fodou 1 Dandou Baïdou 1 Kayla Kama Sagnola Dabadou Blassana Kamian Laye Kondiadou Tignèko Kovila Komende Kassadou Solomana Bengoua Poveni Malla Angola Sokodou Niansoumandou Diani District Capital Kokouma Nongoa Koïko Frandou Sinko Ferela Bolodou Famoîla Mandou Moya Koya Nafadji Domba Koberno Mano Kama Baïzéa Vassala Madina Sèmèkoura Bagbé Yendemillimo Kambadou Mohomè Foomè Sondou Diaboîdou Malondou Dabadou Otol Beindou Koindou -
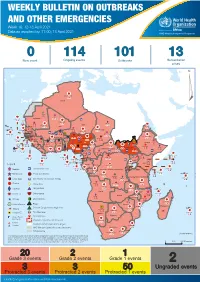
Week 16: 12-18 April 2021
WEEKLY BULLETIN ON OUTBREAKS AND OTHER EMERGENCIES Week 16: 12-18 April 2021 Data as reported by: 17:00; 18 April 2021 REGIONAL OFFICE FOR Africa WHO Health Emergencies Programme 0 114 101 13 New event Ongoing events Outbreaks Humanitarian crises 119 642 3 155 Algeria ¤ 36 13 110 0 5 694 170 Mauritania 7 2 13 070 433 110 0 7 0 Niger 17 129 453 Mali 3 491 10 567 0 6 0 2 079 4 4 706 169 Eritrea Cape Verde 39 782 1 091 Chad Senegal 5 074 189 61 0 Gambia 27 0 3 0 20 466 191 973 5 Guinea-Bissau 847 17 7 0 Burkina Faso 236 49 242 028 3 370 0 164 233 2 061 Guinea 13 129 154 12 38 397 1 3 712 66 1 1 23 12 Benin 30 0 Nigeria 1 873 72 0 Ethiopia 540 2 481 5 6 188 15 Sierra Leone Togo 3 473 296 61 731 919 52 14 Ghana 5 787 75 Côte d'Ivoire 10 473 114 14 484 479 63 0 40 0 Liberia 17 0 South Sudan Central African Republic 916 2 45 0 97 17 25 0 21 612 260 45 560 274 91 709 771 Cameroon 7 0 28 676 137 5 330 13 151 653 2 481 655 2 43 0 119 12 6 1 488 6 4 028 79 12 533 7 259 106 Equatorial Guinea Uganda 542 8 Sao Tome and Principe 32 11 2 066 85 41 378 338 Kenya Legend 7 611 95 Gabon Congo 2 012 73 Rwanda Humanitarian crisis 2 275 35 23 888 325 Measles 21 858 133 Democratic Republic of the Congo 10 084 137 Burundi 3 612 6 Monkeypox Ebola virus disease Seychelles 28 956 745 235 0 420 29 United Republic of Tanzania Lassa fever Skin disease of unknown etiology 190 0 4875 25 509 21 Cholera Yellow fever 1 349 5 6 257 229 24 389 561 cVDPV2 Dengue fever 90 918 1 235 Comoros Angola Malawi COVID-19 Chikungunya 33 941 1 138 862 0 3 815 146 Zambia 133 0 Mozambique -

Observing the 2010 Presidential Elections in Guinea
Observing the 2010 Presidential Elections in Guinea Final Report Waging Peace. Fighting Disease. Building Hope. Map of Guinea1 1 For the purposes of this report, we will be using the following names for the regions of Guinea: Upper Guinea, Middle Guinea, Lower Guinea, and the Forest Region. Observing the 2010 Presidential Elections in Guinea Final Report One Copenhill 453 Freedom Parkway Atlanta, GA 30307 (404) 420-5188 Fax (404) 420-5196 www.cartercenter.org The Carter Center Contents Foreword ..................................1 Proxy Voting and Participation of Executive Summary .........................2 Marginalized Groups ......................43 The Carter Center Election Access for Domestic Observers and Observation Mission in Guinea ...............5 Party Representatives ......................44 The Story of the Guinean Security ................................45 Presidential Elections ........................8 Closing and Counting ......................46 Electoral History and Political Background Tabulation .............................48 Before 2008 ..............................8 Election Dispute Resolution and the From the CNDD Regime to the Results Process ...........................51 Transition Period ..........................9 Disputes Regarding First-Round Results ........53 Chronology of the First and Disputes Regarding Second-Round Results ......54 Second Rounds ...........................10 Conclusion and Recommendations for Electoral Institutions and the Framework for the Future Elections ...........................57 -

Improving Sustainable Water Access in Rural Communities in Guinea
© CEAD 2016 IMPROVING SUSTAINABLE WATER ACCESS IN RURAL COMMUNITIES IN GUINEA OCTOBER 2018 Overview The Ebola outbreak of 2014-2016 highlighted the need to strengthen water, sanitation and hygiene (WASH) health-related preventive measures at all levels in Guinea. Safe drinking water and adequate sanitation are essential for life and health. A sustainable supply of safe water, adequate sanitation and improved hygiene not only saves lives, but also has significant effects for girls and women, who bear primary responsibility for fetching water, which is often unclean and far from home. This daily chore exposes them to the potential risk of violence and can prevent girls from attending school. UNICEF believes that access to WASH goes far beyond health improvements, positively affecting areas such as human rights, girls’ education, gender relations and nutrition. In 2000, approximately 47 percent of the population in rural Guinea consumed contaminated water, further increasing the risk of communicable diseases; the success of the manual drilling project supported by your previous gift has helped bring that percentage down by 2015 to 32 percent.1 Still, rural communities remain poorly served with too few sources of safe water in areas that are too widely spaced apart. UNICEF estimates that at approximately 10,154 water points are required to meet to the country’s basic water supply needs. And although the number of people sharing a water point should only be 300, data collected by the national water service agency (Service National d’Aménagement des Points d’Eau, or SNAPE) shows that an average of 1,500 people in fact share the same water point. -

Elements De La Carte Sanitaire Des Etablissements De Soins Du Secteur Public
REPUBLIQUE DE GUINEE ----------------- Travail - Justice - Solidarité MINISTERE DE LA SANTE PUBLIQUE ELEMENTS DE LA CARTE SANITAIRE DES ETABLISSEMENTS DE SOINS DU SECTEUR PUBLIC JANVIER 2012 TABLE DES MATIERES CHAPITRE I : SITUATION ACTUELLE DES ETABL ISSEMENTS DE SOINS ............................................. 4 I - TYPOLOGIE ............................................................................................................................................................. 4 II - NIVEAU PRIMAIRE ............................................................................................................................................ 4 III - NIVEAU SECONDAIRE ..................................................................................................................................... 7 IV - NIVEAU TERTIAIRE ........................................................................................................................................ 10 V - POINTS FAIBLES .............................................................................................................................................. 12 VI - POINTS FORTS ................................................................................................................................................ 13 VII - CONTRAINTES ............................................................................................................................................... 13 VIII - OPPORTUNITES ET MENACES .................................................................................................................. -

Non-Wood Forest Products and Services for Socio-Economic Development
Non-Wood Forest Products and Services for Socio-Economic Development The AFRICAN FOREST FORUM – a platform for stakeholders in African Forestry A Compendium for Technical and Professional Forestry Education Suzana Agustino, Bennet Mataya, Kingiri Senelwa & Enoch G. Achigan-Dako Non-Wood Forest Products and Services for Socio-Economic Development © African Forest Forum 2011 All rights reserved African Forest Forum c/o World Agroforestry Centre (ICRAF) P.O. Box 30677-00100 Nairobi KENYA Tel: +254 20 722 4203 Fax: +254 20 722 4001 www.afforum.org Disclaimer The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the African Forest Forum concerning the legal status of any country, territory, city or area or its authorities, or concerning the delimitation of its frontiers or boundaries regarding its economic system or degree of development. Excerpts may be reproduced without authorization, on condition that the source is indicated. Views expressed in this publication do not necessarily reflect those of the African Forest Forum. Correct citation: Agustino, S., Mataya, B., Senelwa, K., and Achigan-Dako, G.E. 2011. Non-wood forest products and services for socio-economic development. A Compendium for Technical and Professional Forestry Education. The African Forest Forum, Nairobi, Kenya. 219 pp. Printed in Nairobi in 2011. ISBN 978-92-9059-294-5. Non-Wood Forest Products and Services for Socio-Economic Development A Compendium for Technical and Professional Forestry Education Suzana Agustino Bennet Mataya Kingiri Senelwa Enoch G. Achigan-Dako The African Forest Forum (AFF) NON-WOOD FOREST PRODUCTS AND SERVICES FOR ECONOMIC DEVELOPMENT Table of Contents List of Figures, Tables and Boxes vi Acronymns and Abbreviations viii Foreword xi Concepts, Values and Uses of Non-Wood Forest Products and Services 1 1.1. -

Key Considerations: 2021 Outbreak of Ebola in Guinea, the Context of N’Zérékoré
KEY CONSIDERATIONS: 2021 OUTBREAK OF EBOLA IN GUINEA, THE CONTEXT OF N’ZÉRÉKORÉ This brief summarises key considerations about the social, political and economic context shaping the outbreak of Ebola in the N’Zérékoré prefecture, Guinea, as of March 2021. The outbreak was declared on 14 February 2021, two weeks after the death of the first known case, a health agent (Agent Technique de Santé) from Gouécké. Gouécké is located 40km north of N’Zérékoré via the paved Route Nationale 2. The nurse sought care at a health centre in Gouécké, a clinic and then a traditional healer in N’Zérékoré. She died in N’Zérékoré on 28 January.1 When they became sick, the relatives of the first known case referred themselves to N’Zérékoré regional hospital, where the disease was transmitted to healthcare workers. Although the potential for transmission in rural areas of the Gouécké subprefecture was high, to date, most cases have been reported in the urban setting of N’Zérékoré, which is the focus of this brief. At the time of writing (22 March), the total number of cases was 18 (14 confirmed, four probable), with nine deaths and six recoveries. The last new case was reported on 4 March. N’Zérékoré city is the administrative capital of N’Zérékoré prefecture and N’Zérékoré region, and the largest urban centre of the Guinée Forestière area of south-east Guinea. It has an estimated population of 250,000, and is an important commercial, economic and transportation hub connecting Guinée Forestière to neighbouring Sierra Leone, Liberia and Côte d’Ivoire. -

Rapport-Final-UFDG.Pdf
UFDG Union des Forces Démocratiques de Guinée ELECTION PRESIDENTIELLE DU 18 OTCOBRE 2020 RAPPORT SYNTHESE DE L’ÉLECTION PRÉSIDENTIELLE DU 18 OCTOBRE 2020 1 UFDG Union des Forces Démocratiques de Guinée TABLE DE MATIERE Résumé …………………………………………………………………………………………………………………………………………3 Partie 1 : Présentation des résultats de l’élection présidentielle du 18 octobre 2020 suivant le dispositif de comptabilisation des résultats par l’UFDG …………………………………………………………........5 Partie 2 : Mise en exergue du système de falsification des résultats dans la comptabilisation des voix effectuée par la CENI …………………………………………………………………………………………………………….9 2.1 : Incohérences sur les taux de participation et la sincérité du vote dans certaines circonscriptions électorales de la haute guinée ……………………………………………………………………..…….9 2.2 : Bourrages effectués sur des bureaux de vote de moins de 100 électeurs et pour lesquels les nombres de votants enregistrés le jour du scrutin sont largement supérieurs aux nombres d’inscrits principalement dans les fiefs du RPG …………………………………………………………………………..35 2.3 : Bureaux de vote écartés par les commissions administratives de centralisation (CACV) ………………………………………………………………………………………………………………………………………………….…38 2.4: Falsifications des résultats dans les CACV en faveur du RPG ……………………………………………....43 2 UFDG Union des Forces Démocratiques de Guinée RESUME Les résultats provisoires de l’élection présidentielle du 18 octobre 2020 proclamés par la CENI prouvent à suffisance les limites de cette structure dans la conduite du processus électoral afin d’obtenir des résultats techniques tangibles, crédibles et acceptables par toutes les parties prenantes. Les partenaires techniques et financiers, les ONG et les observateurs déployés sur le terrain ont tous relayé des disfonctionnements majeurs enregistrés le jour du scrutin en mettant un accent particulier sur les incohérences des chiffres publiés par la CENI, et la transformation des CACV en centre de modification des résultats. -

Rapport Synthese De L'élection Présidentielle Du
UFDG Union des Forces Démocratiques de Guinée ELECTION PRESIDENTIELLE DU 18 OTCOBRE 2020 RAPPORT SYNTHESE DE L’ÉLECTION PRÉSIDENTIELLE DU 18 OCTOBRE 2020 1 UFDG Union des Forces Démocratiques de Guinée TABLE DE MATIERE Résumé …………………………………………………………………………………………………………………………………………3 Partie 1 : Présentation des résultats de l’élection présidentielle du 18 octobre 2020 suivant le dispositif de comptabilisation des résultats par l’UFDG …………………………………………………………........5 Partie 2 : Mise en exergue du système de falsification des résultats dans la comptabilisation des voix effectuée par la CENI …………………………………………………………………………………………………………….9 2.1 : Incohérences sur les taux de participation et la sincérité du vote dans certaines circonscriptions électorales de la haute guinée ……………………………………………………………………..…….9 2.2 : Bourrages effectués sur des bureaux de vote de moins de 100 électeurs et pour lesquels les nombres de votants enregistrés le jour du scrutin sont largement supérieurs aux nombres d’inscrits principalement dans les fiefs du RPG …………………………………………………………………………..35 2.3 : Bureaux de vote écartés par les commissions administratives de centralisation (CACV) ………………………………………………………………………………………………………………………………………………….…38 2.4: Falsifications des résultats dans les CACV en faveur du RPG ……………………………………………....43 2 UFDG Union des Forces Démocratiques de Guinée RESUME Les résultats provisoires de l’élection présidentielle du 18 octobre 2020 proclamés par la CENI prouvent à suffisance les limites de cette structure dans la conduite du processus électoral afin d’obtenir des résultats techniques tangibles, crédibles et acceptables par toutes les parties prenantes. Les partenaires techniques et financiers, les ONG et les observateurs déployés sur le terrain ont tous relayé des disfonctionnements majeurs enregistrés le jour du scrutin en mettant un accent particulier sur les incohérences des chiffres publiés par la CENI, et la transformation des CACV en centre de modification des résultats. -

Ebola Situation Report
UNICEF Guinea | Ebola Situation Report EBOLA Situation Report #7 14 May – 1st June 2021 Epidemiological Situation st (as of 1 June 2021) @UNICEF Guinea/Saa Momory Koundouno 16 confirmed cases Situation Overview 5 deaths • As of 1st June 2021, 16 cases of Ebola were confirmed in Guinea, including 5 confirmed deaths and 10 recoveries. • 10,873 people have been inoculated (including 2,879 frontline staff) 10 people recovered since the official launch of the vaccination on February 23, 2021. • The countdown (42 days) to the end of the epidemic started on May 8, 2021. Guinea is 19 days away from declaring the end of the Ebola virus 1 region affected disease. • 10,120 people directly sensitized (including 4,539 women) through preaching and sermons by imams and priests in 30 mosques and 20 churches on preventive measures against Ebola in the urban commune of N'Zérékoré. • 60 humanitarian actors trained on the prevention of sexual exploitation and abuse, the survivor-centered approach for the management of GBV and community- based complaint mechanisms. • The funding gap for the response is currently at 81%. UNICEF Guinea | Ebola Situation Report 14 May – 1st June 2021 UNICEF Guinea| Ebola Situation 15 February – 10 March 21 Report February 2021 UNICEF Response Infection Prevention and Control (IPC) and Water, Hygiene and Sanitation (WASH) UNICEF continued to distribute hygiene kits, reinforce water supply mechanism and sanitation in health facilities and in the community. • 55 hygiene kits have been distributed to social mobilizers in Boma in the urban commune of N’Zérékoré. • Finalization of the rehabilitation works and installation of solar equipment for the well in the epidemiological treatment center of Gouécké. -

Guinea: Reference Map of N'zérékoré Prefecture
Guinea:ReferenceReference MapMap ofof NZérékoréN’Zérékoré Prefecture Prefecture(as (as of of 2612 MarchFev 2015)2015) Boola Mazana Kpamou Ouota LÖMOU CENTRE GOUNAGALAYE CENTRE KONI CENTRE Kognea Saadou Kpeïda MAKPOZOU CENTRE Founbandou Lokoa Koropara Lamineta FOOZOU CENTRE BOMA-NORD CENTRE Yessila Gbana Bahaïta Ouleoun KÔRÔH CENTRE KABIETA Kelèta KOLIWATA CENTRE Suokpayai PAMPARA CENTRE NYEMA-NORD CENTRE Gbohileta Bhoita BOWE NORD TOKPATA CENTRE Laine Zebela KEORAH CENTRE Womey KEREZAGAYE CENTRE Beneouli Göta ZENEMOUD CENTRE Nyonta GBEANTA CENTRE Seloh Loula KOYAH CENTRE Koule OUEYE CENTRE YOMATA CENTRE Oulela KOMATA CENTRE Bapet GOUECKE CENTRE Gouecke Vomou TAMOE CENTRE PAMPORE CENTRE Gnanakpalaye KPAI CENTRE Nyineta Guelagbozou BANZOU CENTRE KPOULO CENTRE ZOGBEANTA CENTRE Dapoloye Kpaya Gbonoma Yarakpara Soulouta Konia Benoita KPAGALAYE CENTRE Youwa NONA CENTRE MAOUON CENTRE KEREMANDA CENTRE Koronta GBOUO CENTRE ZOGOTA CENTRE Gbagonaye Belefanaye Koliyapoulou Powai SOUHOULE centre Kokota WEYA SUD Gouh GBAMBA CENTRE NIENH CENTRE Kpatié Yelewena Kobela NIAMPARA CENTRE Teaye Banzou Sud Kpeliyewoun 2 Batouata Woroyé Kpeliyewoun 1 NYEMA SUD CENTRE KONIPARA CENTRE Gbaita Duala GBILI CENTRE Nyalakpale GALAKPAYE CENTRE Ninguekpoye KOMOU CENTRE Weya Nord Domou 2 Samoi Domou 1 GBAYA CENTRE Lola KONIA AVIATION CENTRE Guela Boma Sud Bowe LOULE NORD KONALA CENTRE GBOTTOYE CENTRE MOATA CENTRE KOULE SUD CENTRE BANGOUETA CENTRE Galaye NYENSOKOURA 2 KARANA CENTRE KOLEYEBA NYEN 1 GONIA 3 BELLE VUE Zogoya MOHOMOU Lamou Wezou GBOYEBA Nzerekore NZAO CENTRE