Assessing Seed Desiccation Responses of Native Trees in the Caribbean
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Natural Resources Conservation Service Conservation General Specifications
NATURAL RESOURCES CONSERVATION SERVICE CONSERVATION GENERAL SPECIFICATIONS RIPARIAN FOREST BUFFER (Acre) Code 391 Procedures, technical details and other establishing new buffers. Ecological life information listed below provide additional zones, described by Ewel & Withmore, 1973, guidance for carrying out selected for Puerto Rico and the U. S. Virgin Islands components of the Riparian Forest Buffer. are major climatic divisions and define the This material is referenced from the conditions for ecosystem functioning (see conservation practice standard for the pages 5 and 6 for Ecological Life Zones map named practice and supplements the for Puerto Rico and U. S. Virgin Islands). requirements and considerations listed therein. Natural regeneration may be used to establish a buffer if the following conditions PLANTING DENSITIES exist: Initial plant-to-plant densities for trees and • There is an adequate natural seed shrubs will depend on their potential height source of desired species in adjacent at 20 years of age. Heights may be areas. estimated based on performance of the • Site conditions are favorable for individual species (or comparable species) establishing the desired number and in nearby areas on similar sites, or distribution of seedlings within specified technical references. Planting density time period. specifications are: • Noxious or invasive species are not likely to jeopardize the stand. Plant Plant-to-Plant CARE, HANDLING, SIZE AND PLANTING Types/Heights: Spacing (ft) REQUIREMENTS FOR WOODY PLANTING Shrubs 3 to 15 STOCK Trees 8 to 20 Planting stock will be stored in a cool, moist environment. During all stages of handling PLANT LIST and storage, keep stock tops dry and free of Table 1 includes the lists of woody plant mold and roots moist and cool. -

Samara Newsletter July & August 2020
SamaraThe International Newsletter of the Millennium Seed Bank Partnership Special issue featuring projects and research from The Global Tree Seed Bank Programme, funded by the Garfield Weston Foundation August/September 2020 Issue 35 ISSN 1475-8245 Juglans pyriformis in the State of Veracruz Conserving and investigating native tree seeds to support community-based reforestation initiatives in Mexico Veracruz Pronatura Photo: Mexico is the fourth richest country in the world in terms of plant Millennium Seed Bank. Seed research has species diversity, after Brazil, China, and Colombia with a flora of been carried out on 314 species to study ca. 23,000 vascular plants. Around half of the plant species are their tolerance to desiccation for seed endemic and nearly 3,500 are trees. banking and to determine germination requirements to inform propagation activities. One of the key project species ELENA CASTILLO-LORENZO (Latin America Projects Coordinator, RBG Kew), MICHAEL WAY is Cedrela odorata (Spanish cedar), whose (Conservation Partnership Coordinator (Americas, RBG Kew) & TIZIANA ULIAN (Senior Research conservation status is vulnerable (IUCN Leader – Diversity and Livelihoods, RBG Kew) 2020) due to exploitation for its highly Trees and forests provide multiple goods Iztacala of the Universidad Autónoma valued wood. C. odorata is also used for and benefits for humans, such as high- de México (Fes-I UNAM). The aim medicinal purposes by local communities quality wood, fruit, honey, and other of this project was to conserve tree in Mexico, with the leaves being prepared ecosystem services, including clean water, species through a collaborative research in herbal tea to treat toothache, earache, prevention of soil erosion and mitigation of programme focusing on endemic, and intestinal infections. -

Caribbean Ornithology
The Journal of Caribbean Ornithology RESEARCH ARTICLE Vol. 29:21–27. 2016 Avian biodiversity in a pasture-dominated ecosystem Jason P. Hernandez Photo: J.P. Hernandez The Journal of Caribbean Ornithology www.birdscaribbean.org/jco ISSN 1544-4953 RESEARCH ARTICLE Vol. 29:21–27. 2016 www.birdscaribbean.org Avian biodiversity in a pasture-dominated ecosystem Jason P. Hernandez Abstract Burgeoning human populations have created many challenges for conservation of biodiversity, as ever-larger areas of land are converted to agricultural and other forms of production. Protected reserves alone are not sufficient to sustain biodiversity in a world of increasing human needs, so it is necessary to understand the extent to which biodiversity can exist with- in highly altered, agricultural ecosystems. From 11 May to 8 August 2013, bird diversity was surveyed at an agricultural site on Hispaniola, utilized mainly for pasture, and the population and regeneration potential of the royal palm (Roystonea borinquena), a key nest tree species for two abundant bird species, was assessed. The site harbored 33 bird species including 32 resident species (about one-third of Hispaniola’s low-elevation resident land bird species), of which 5 are endemic to Hispaniola. Two of these endemic species, the Palmchat (Dulus dominicus) and the Hispaniolan Woodpecker (Melanerpes striatus), were the pre- dominant species present. Seventy-six percent of species at this site showed an association with trees. The royal palm occurred mainly in wooded riparian corridors, but showed little ability to colonize pastures, suggesting possible declines in future overall tree density at this site. Ensuring adequate tree recruitment is crucial to maintaining biodiversity in agricultural and grazing zones. -

Monocotyledons and Gymnosperms of Puerto Rico and the Virgin Islands
SMITHSONIAN INSTITUTION Contributions from the United States National Herbarium Volume 52: 1-415 Monocotyledons and Gymnosperms of Puerto Rico and the Virgin Islands Editors Pedro Acevedo-Rodríguez and Mark T. Strong Department of Botany National Museum of Natural History Washington, DC 2005 ABSTRACT Acevedo-Rodríguez, Pedro and Mark T. Strong. Monocots and Gymnosperms of Puerto Rico and the Virgin Islands. Contributions from the United States National Herbarium, volume 52: 415 pages (including 65 figures). The present treatment constitutes an updated revision for the monocotyledon and gymnosperm flora (excluding Orchidaceae and Poaceae) for the biogeographical region of Puerto Rico (including all islets and islands) and the Virgin Islands. With this contribution, we fill the last major gap in the flora of this region, since the dicotyledons have been previously revised. This volume recognizes 33 families, 118 genera, and 349 species of Monocots (excluding the Orchidaceae and Poaceae) and three families, three genera, and six species of gymnosperms. The Poaceae with an estimated 89 genera and 265 species, will be published in a separate volume at a later date. When Ackerman’s (1995) treatment of orchids (65 genera and 145 species) and the Poaceae are added to our account of monocots, the new total rises to 35 families, 272 genera and 759 species. The differences in number from Britton’s and Wilson’s (1926) treatment is attributed to changes in families, generic and species concepts, recent introductions, naturalization of introduced species and cultivars, exclusion of cultivated plants, misdeterminations, and discoveries of new taxa or new distributional records during the last seven decades. -

Sfps Fall 2011 Sale Plant List
SFPS FALL 2011 SALE PLANT LIST PLANTS VENDOR # Palms Acanthophoenix rubra 35 Acoelorrhaphe wrightii 26, 67 Acrocomia aculeata 50, 67 Actinokentia divaricata 35, 57, 66, 68, 72 Actinorhytis calapparia 72 Adonidia merrillii 31, 57, 66, 89 Adonidia merrillii var. "Golden Form" 35 Aiphanes aculeata = Aiphanes horrida - Aiphanes caryotifolia = Aiphanes horrida - Aiphanes erosa = Aiphanes minima - Aiphanes horrida 35, 68, 72 Aiphanes minima 68 Aiphanes vincentiana = Aiphanes minima - Allagoptera arenaria 57, 66, 67, 68, 72 Allagoptera campestris 67 Allagoptera leucocalyx 57 Alloschmidia glabrata = Basselinia glabrata - Alsmithia longipes = Heterospathe longipes - Archontophoenix cunninghamiana var. 'Illawara' 68 Archontophoenix maxima 67, 72 Archontophoenix myolensis 50, 66, 67, 68 Archontophoenix purpurea 57, 66, 72 Archontophoenix tuckeri 66, 68 Areca aliceae = Areca triandra - Areca camarinensis 57, 68 Areca catechu 57, 67, 72 Areca catechu var. 'Dwarf' 35, 50 Areca hutchinsoniana 68 Areca ipot 67 Areca latiloba = Areca montana - Areca macrocalyx var. 'Red Form' 35, 57, 68 Areca macrocarpa 68 Areca montana 57 Areca triandra 68, 72 Areca vestiaria 25, 35, 57, 67, 68 Areca vestiaria var. 'Orange Form' 25, 57, 67, 72 Areca vestiaria var. 'Maroon Leaf' 35, 57, 67 Areca vestiaria var. 'Red Leaf' 57, 67, 72 Areca sp. 'Yellow Crownshaft' 25 Arenga ambong = Arenga undulatifolia - Arenga brevipes 57 Arenga caudata 66 Arenga engleri 31, 66, 68, 72 Arenga hookeriana 35, 57, 66, 72 Arenga microcarpa 26, 66 Arenga obtusifolia 57, 66 PLANTS VENDOR # Arenga pinnata 50, 57, 66, 67, 68 Arenga porphyrocarpa 66 Arenga tremula 26, 57, 66, 68, 72 Arenga undulatifolia 35, 57, 66, 67 Arenga westerhoutii 68 Asterogyne martiana 57, 68, 72 Astrocaryum acaule 72 Astrocaryum alatum 35, 50, 57, 67 Astrocaryum mexicanum 72 Astrocaryum murumuru 72 Attalea butyracea 57, 67, 72 Attalea cohune 35 Attalea phalerata 50, 91 Attalea rostrata 68 Attalea speciosa 50, 66 Bactris bidentula 72 Bactris gasipaes 67 Bactris gasipaes var. -
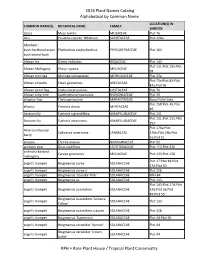
2016 Plant Names Catalog Alphabetical by Common Name
2016 Plant Names Catalog Alphabetical by Common Name LOCATION(S) IN COMMON NAME(S) BOTANICAL NAME FAMILY GARDEN abaca Musa textilis MUSACEAE Plot 76 abiu Pouteria caimito 'Whitman' SAPOTACEAE Plot 128a Abraham- bush:hardhead:scipio- Phyllanthus epiphyllanthus PHYLLANTHACEAE Plot 164 bush:sword-bush African iris Dietes iridioides IRIDACEAE Plot 143 Plot 131:Plot 19a:Plot African Mahogany Khaya nyasica MELIACEAE 58 African moringa Moringa stenopetala MORINGACEAE Plot 32a Plot 71a:Plot 83:Plot African oil palm Elaeis guineensis ARECACEAE 84a:Plot 96 African spiral flag Costus lucanusianus COSTACEAE Plot 76 African tulip-tree Spathodea campanulata BIGNONIACEAE Plot 29 alligator flag Thalia geniculata MARANTACEAE Royal Palm Lake Plot 158:Plot 45:Plot allspice Pimenta dioica MYRTACEAE 46 Amazon lily Eucharis x grandiflora AMARYLLIDACEAE Plot 131 Plot 131:Plot 151:Plot Amazon-lily Eucharis amazonica AMARYLLIDACEAE 152 Plot 176a:Plot American beauty Callicarpa americana LAMIACEAE 176b:Plot 19b:Plot berry 3a:Plot 51 anaqua Ehretia anacua BORAGINACEAE Plot 52 anchovy pear Grias cauliflora LECYTHIDACEAE Plot 112:Plot 32b andiroba:bastard Carapa guianensis MELIACEAE Plot 133:Plot 158 mahogany Plot 17:Plot 18:Plot angel's trumpet Brugmansia aurea SOLANACEAE 27d:Plot 50 angel's trumpet Brugmansia aurea x SOLANACEAE Plot 32b angel's trumpet Brugmansia 'Ecuador Pink' SOLANACEAE RPH-B4 angel's trumpet Brugmansia sp. SOLANACEAE Plot 133 Plot 143:Plot 27d:Plot angel's trumpet Brugmansia suaveolens SOLANACEAE 32b:Plot 3a:Plot 49:Plot 50 Brugmansia suaveolens -

ABSTRACT the Endangered Puerto Rican Parrot (Amazona Vittata) Has
ABSTRACT MALONE, MEREDITH BROOKE. Abundance, Fruit Presence, and Growth Enhancement of Prestoea montana and Roystonea borinquena, Two Palm Species of Importance to Amazona vittata (Puerto Rican Parrot), in Río Abajo Forest, Puerto Rico. (Under the direction of Daniel J. Robison.) The endangered Puerto Rican parrot (Amazona vittata) has been bred in captivity for decades, and since 2000, over 50 parrots have been released into native habitats to rebuild wild populations in Puerto Rico. One group of 21 parrots was released into Río Abajo Forest in the karstic region of Puerto Rico in November 2006, where high quality habitat resources exist, but the management of sustainable food resources, such as the fruit of Prestoea montana (sierra palm) and Roystonea borinquena (royal palm), is imperative. An inventory of accessible regions in Río Abajo Forest was conducted to assess differences in size and abundance of P. montana and R. borinquena palms between three forest community types and fruiting patterns under differing growing conditions. Twenty palms in each of three size classes of both palm species were located in Río Abajo Forest and approximately half were supplied with a controlled-release formula of inorganic NPK fertilizer with micronutrients to investigate potential soil nutrient limitations to growth of palms. Thirty R. borinquena seedlings were located in artificial canopy gap plots previously created in abandoned Hibiscus elatus (mahoe) plantations in Río Abajo Forest and approximately half were supplied with the same fertilizer formula to evaluate combined soil and light limitations on seedling growth. In the size and abundance inventory, seedlings were the most abundant size class of both palm species. -

Hort Pro Version V List For
HORTICOPIA® Professional Woody Plus Refresh Library Plant List Name Name Abelia 'Mardi Gras' Acalypha wilkesiana 'Petticoat' Abelia x grandiflora 'John Creech' Acer buergerianum 'Goshiki kaede' Abelia x grandiflora 'Sunshine Daydream' Acer campestre 'Carnival' Abelia schumannii 'Bumblebee' Acer campestre 'Evelyn (Queen Elizabeth™)' Abies concolor 'Compacta' Acer campestre 'Postelense' Abies concolor 'Violacea' Acer campestre 'Tauricum' Abies holophylla Acer campestre var. austriacum Abies koreana 'Compact Dwarf' Acer cissifolium ssp. henryi Abies koreana 'Prostrate Beauty' Acer davidii ssp. grosseri Abies koreana 'Silberlocke' Acer elegantulum Abies nordmanniana 'Lowry' Acer x freemanii 'Armstrong II' Abies nordmanniana 'Tortifolia' Acer x freemanii 'Celzam' Abies pindrow Acer x freemanii 'Landsburg (Firedance®)' Abies pinsapo 'Glauca' Acer x freemanii 'Marmo' Abies sachalinensis Acer x freemanii 'Morgan' Abutilon pictum 'Aureo-maculatum' Acer x freemanii 'Scarlet Sentenial™' Acacia albida Acer heldreichii Acacia cavenia Acer hyrcanum Acacia coriacea Acer mandschuricum Acacia erioloba Acer maximowiczianum Acacia estrophiolata Acer miyabei 'Morton (State Street®)' Acacia floribunda Acer mono Acacia galpinii Acer mono f. dissectum Acacia gerrardii Acer mono ssp. okamotoanum Acacia graffiana Acer monspessulanum Acacia karroo Acer monspessulanum var. ibericum Acacia nigricans Acer negundo 'Aureo-marginata' Acacia nilotica Acer negundo 'Sensation' Acacia peuce Acer negundo 'Variegatum' Acacia polyacantha Acer oliverianum Acacia pubescens Acer -

Introduced Palm Flora– 5
INTRODUCED PALM FLORA– 5 5.1. Introduction Palms are one of the most attractive well-known, highly cultivated economically most important plants in the world. It has important roles to the human throughout much of history. Introduced palms are much widely used in landscaping for their beautiful appearances (Roystonea regia, Roystonea borinquena, Raphis excelsa, R. humilis, Ptychosperma macarthurii, Chamedorea elegans etc.) and sometimes as cultivated crops (Elaeis guineensis). There were about 2600 species of ornamental palms that restricted to the various climatic zones of tropical, subtropical to humid regions of the world. In India as well as in West Bengal it is not exactly known about the exact numbers of exotic species of palms because quite a large number of them introduced by private growers. In some important government and public gardens in West Bengal palms are seldom labelled and listed with their correct nomenclature and thus case inconvenience to the palm lovers (Basu and Chakraverty 1994). Introduction and cultivation of exotic palms in India began in the early 18th century when Arabian date palm (Phoenix dactylifera) was brought for cultivation. Cultivation of indigenous and exotic palms on a scientific basis was started after the establishment of East India Company’s garden in West Bengal, when Malayan Sago palm Metroxylon sagu was introduced to India. Latter on several palms like Indonesian sugar palm (Arenga pinnata), African oil palm (Elaeis guineensis) South American oil yielding Babasunut palm (Orbignya cohune) were also introduced. Several exotic species have been introduced and many of them are cultivated at present as ornamental palms in the various gardens (Renuka 2010). -

Structure and Species Composition of Novel Forests Dominated by an Introduced Species in Northcentral Puerto Rico
New Forests (2010) 39:1–18 DOI 10.1007/s11056-009-9154-7 Structure and species composition of novel forests dominated by an introduced species in northcentral Puerto Rico Oscar J. Abelleira Martı´nez Æ Mariela A. Rodrı´guez Æ Ivonne Rosario Æ Nataly Soto Æ Armando Lo´pez Æ Ariel E. Lugo Received: 9 January 2009 / Accepted: 9 May 2009 / Published online: 27 May 2009 Ó Springer Science+Business Media B.V. 2009 Abstract The African tulip tree, Spathodea campanulata Beauv., is an introduced spe- cies forming novel forest types in Puerto Rico. These forests develop naturally after deforestation, agricultural use and land abandonment, and there are many questions as to their ecological characteristics. We sampled structure and species composition of large, small, and juvenile trees (C10, C2.5 to \10, and \2.5 cm diameter at breast height, respectively) in nine secondary forests dominated by S. campanulata on alluvial, karst, and volcanic substrates in northcentral Puerto Rico. No differences were found in S. cam- panulata forest structure between substrates. Of a total of 79 species found, 17 were introduced. Forests on karst and alluvium had the highest and lowest global species richness, respectively. Species richness increased from large to small to juvenile trees in most sites, but more so on karst. The percentage of introduced species was inversely related to species richness of tree size classes on all substrate types. The dominance of S. campanulata in the large tree size class was highest and lowest in alluvial and volcanic sites, respectively, and decreased from large to small to juvenile trees on all substrate types. -

Index to Volume 49 Vol
PALMS Index to volume 49 Vol. 49(4) 2005 Calamus heteroideus 26–29 Index to Volume 49 Calamus horrens 30 Calamus javensis 26–29, 34 A journey along the Rio Napo 85 Calamus kingianus 118 A key to common landscape palm disorders and Calamus leptospadix 118 diseases in the continental United States 143 Calamus melanoloma 26–30 A little palm, a lot of palaver 48 Calamus nambariensis 118, 156 A new cultivar of Sabal palmetto 46 Calamus occidentalis 30 Acrocomia 184 Calamus optimus 31 Acocomia aculeata 185 Calamus ornatus 26–30, 33, 34 Adams, B., as co-author 48 Calamus palustris 118 Adonidia merrillii 147 Calamus polystachys 26–29, 34 Allagoptera 122 Calamus rhomboideus 26–30, 34 Archontophoenix cunninghamiana 192, 194 Calamus tenuis 116, 118 Areca triandra 118 Calamus trachycoleus 31 Arenga 6, 7, 8, 31 Calamus viminalis 117, 120 Arenga engleri 192 Calyptrogyne 149 Arenga obtusifolia 6–14 Calyptrogyne occidentalis 149 Arenga pinnata 3, 5–12, 14, 25, 31, 52 Calyptrogyne plumeriana 149 Arenga westerhoutii 6–14, 118 Calyptrogyne plumeriana, a new name for a familiar Astrocaryum 90 palm 149 Astrocaryum chambira 90 Calyptrogyne rivalis 149 Astrocaryum jauari 89, 90 Calyptronoma 149 Astrocaryum macrocalyx 87, 89, 90 Calyptronoma plumeriana 149 Astrocaryum urostachys 87, 89, 90 Calyptronoma rivalis 185 Attalea 89, 90 Carpentaria acuminata 168, 169, 176, 178, 180 Attalea butyracea 87–90 Caryota 80–82 Attalea crassispatha 185 Caryota “himalayana” 192 Attalea ferruginea 87 Caryota maxima 118 Averyanov, L.V., N.T. Hiep and P.K. Loc: Guihaia Caryota ophiopellis -

Raoiella Indica Global Invasive Species Database (GISD)
FULL ACCOUNT FOR: Raoiella indica Raoiella indica System: Terrestrial Kingdom Phylum Class Order Family Animalia Arthropoda Arachnida Acariformes Tenuipalpidae Common name red palm mite (English), coconut red mite (English), coconut mite (English), red date mite (English), scarlet mite (English), frond crimson mite (English), leaflet false spider mite (English) Synonym Similar species Tetranychus Summary Raoiella indica (the red palm mite) is a parasitic mite invasive in the Caribbean region; it poses a serious threat to many plant industries. Its recent invasion is referred to as the biggest mite explosion in the Americas. Already taking serious tolls on coconut, ornamental palm and orchid crops, its infestation of new species and spread to new locations makes it one the most menacing pests to the Western tropics. view this species on IUCN Red List Species Description Raoiella indica is a bright red mite that parasitizes many important plant species, principally palm and banana species. It can be found on the undersides of their leaves. R. indica resembles the common pests spider mites (Family:Tetranychidae) only with longer spatulate setae and lacking their web spinning ability. Adult females are about 0.32mm long and have dark patches on their abdomen. They are larger than males which have more triangular abdomens. Nymphal stages look similar only smaller with less pronounced setae. The mites are visible with the naked eye and usually congregate in clusters of 100-300, and are surrounded by their white exuvial remains (cast skins). (Kane, 2006; Welbourn, 2007). Lifecycle Stages Eggs are atteched to leaves by a stipe and hatch after an average of 6.5 days.