Phase II Report Chapter 14. Temporal and Geographic Trends in Mercury
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Penobscot Rivershed with Licensed Dischargers and Critical Salmon
0# North West Branch St John T11 R15 WELS T11 R17 WELS T11 R16 WELS T11 R14 WELS T11 R13 WELS T11 R12 WELS T11 R11 WELS T11 R10 WELS T11 R9 WELS T11 R8 WELS Aroostook River Oxbow Smith Farm DamXW St John River T11 R7 WELS Garfield Plt T11 R4 WELS Chapman Ashland Machias River Stream Carry Brook Chemquasabamticook Stream Squa Pan Stream XW Daaquam River XW Whitney Bk Dam Mars Hill Squa Pan Dam Burntland Stream DamXW Westfield Prestile Stream Presque Isle Stream FRESH WAY, INC Allagash River South Branch Machias River Big Ten Twp T10 R16 WELS T10 R15 WELS T10 R14 WELS T10 R13 WELS T10 R12 WELS T10 R11 WELS T10 R10 WELS T10 R9 WELS T10 R8 WELS 0# MARS HILL UTILITY DISTRICT T10 R3 WELS Water District Resevoir Dam T10 R7 WELS T10 R6 WELS Masardis Squapan Twp XW Mars Hill DamXW Mule Brook Penobscot RiverYosungs Lakeh DamXWed0# Southwest Branch St John Blackwater River West Branch Presque Isle Strea Allagash River North Branch Blackwater River East Branch Presque Isle Strea Blaine Churchill Lake DamXW Southwest Branch St John E Twp XW Robinson Dam Prestile Stream S Otter Brook L Saint Croix Stream Cox Patent E with Licensed Dischargers and W Snare Brook T9 R8 WELS 8 T9 R17 WELS T9 R16 WELS T9 R15 WELS T9 R14 WELS 1 T9 R12 WELS T9 R11 WELS T9 R10 WELS T9 R9 WELS Mooseleuk Stream Oxbow Plt R T9 R13 WELS Houlton Brook T9 R7 WELS Aroostook River T9 R4 WELS T9 R3 WELS 9 Chandler Stream Bridgewater T T9 R5 WELS TD R2 WELS Baker Branch Critical UmScolcus Stream lmon Habitat Overlay South Branch Russell Brook Aikens Brook West Branch Umcolcus Steam LaPomkeag Stream West Branch Umcolcus Stream Tie Camp Brook Soper Brook Beaver Brook Munsungan Stream S L T8 R18 WELS T8 R17 WELS T8 R16 WELS T8 R15 WELS T8 R14 WELS Eagle Lake Twp T8 R10 WELS East Branch Howe Brook E Soper Mountain Twp T8 R11 WELS T8 R9 WELS T8 R8 WELS Bloody Brook Saint Croix Stream North Branch Meduxnekeag River W 9 Turner Brook Allagash Stream Millinocket Stream T8 R7 WELS T8 R6 WELS T8 R5 WELS Saint Croix Twp T8 R3 WELS 1 Monticello R Desolation Brook 8 St Francis Brook TC R2 WELS MONTICELLO HOUSING CORP. -

The Following Document Comes to You From
MAINE STATE LEGISLATURE The following document is provided by the LAW AND LEGISLATIVE DIGITAL LIBRARY at the Maine State Law and Legislative Reference Library http://legislature.maine.gov/lawlib Reproduced from scanned originals with text recognition applied (searchable text may contain some errors and/or omissions) ACTS AND RESOLVES AS PASSED BY THE Ninetieth and Ninety-first Legislatures OF THE STATE OF MAINE From April 26, 1941 to April 9, 1943 AND MISCELLANEOUS STATE PAPERS Published by the Revisor of Statutes in accordance with the Resolves of the Legislature approved June 28, 1820, March 18, 1840, March 16, 1842, and Acts approved August 6, 1930 and April 2, 193I. KENNEBEC JOURNAL AUGUSTA, MAINE 1943 PUBLIC LAWS OF THE STATE OF MAINE As Passed by the Ninety-first Legislature 1943 290 TO SIMPLIFY THE INLAND FISHING LAWS CHAP. 256 -Hte ~ ~ -Hte eOt:l:llty ffi' ft*; 4tet s.e]3t:l:ty tfl.a.t mry' ~ !;;llOWR ~ ~ ~ ~ "" hunting: ffi' ftshiRg: Hit;, ffi' "" Hit; ~ mry' ~ ~ ~, ~ ft*; eounty ~ ft8.t rett:l:rRes. ~ "" rC8:S0R8:B~e tffi:re ~ ft*; s.e]38:FtaFe, ~ ~ ffi" 5i:i'ffi 4tet s.e]3uty, ~ 5i:i'ffi ~ a-5 ~ 4eeme ReCCSS8:F)-, ~ ~ ~ ~ ~ ffi'i'El, 4aH ~ eRtitles. 4E; Fe8:50nable fee5 ffi'i'El, C!E]3C::lSCS ~ ft*; sen-ices ffi'i'El, ~ ft*; ffi4s, ~ ~ ~ ~ -Hte tFeasurcr ~ ~ eouRty. BefoFc tfte sffi4 ~ €of' ~ ~ 4ep i:tt;- ~ ffle.t:J:.p 8:s.aitional e1E]3cfisc itt -Hte eM, ~ -Hte ~ ~~' ~, ftc ~ ~ -Hte conseRt ~"" lIiajority ~ -Hte COt:l:fity COfi111'lissioReFs ~ -Hte 5a+4 coufity. Whenever it shall come to the attention of the commis sioner -

I. NATURAL RESOURCES 1. Introduction a Comprehensive Plan Should Provide an Overview of a Town's Natural Resources. These Reso
I. NATURAL RESOURCES 1. Introduction A comprehensive plan should provide an overview of a town's natural resources. These resources are important to the town in several ways. First, they provide critical wildlife and fisheries habitats. Second, inappropriate development in environmentally fragile areas could be costly to the entire town. For example, disruption of natural drainage patterns could increase the chances of flooding. Finally, these resources are an essential part of the town's rural character and help sustain Orland's quality of life. Specifically, this chapter will: a. describe Orland's critical natural and scenic resources; b. predict whether these resources will be threatened by the impacts of future growth and development; and c. assess the effectiveness of existing efforts to protect and preserve these resources. 2. Key Findings and Issues Orland has one bald eagle nest site, according to state records, and one of the few known locations in Maine for the ram's-head lady slipper plant, a member of the orchid family. There are also several high value waterfowl and wading bird habitats in town. Game species such as deer, black bear, and moose can be found in upland areas. Orland's varied landscape of lakes, river valleys, and ridges means that there are many scenic views. While there are no immediate threats to Orland's natural resources, there are also few measures in place to protect these resources over the long run. 3. Public Opinion Survey and Community Workshop Results About 71 percent of survey respondents said that they favored measures to protect open space and wildlife habitat. -

Up and Up: River Herring in Eastern Maine
DOWNEAST FISHERIES PARTNERSHIP 2019 Up and up: River Herring in Eastern Maine PREPARED BY Anne Hayden, Medea Steinman, and Rachel Gorich This report is also available online at: downeastfisheries.org OUR PARTNERS coa.edu downeastinstitute.org mainesalmonrivers.org coastalfisheries.org mcht.org mainefarmlandtrust.org seagrant.umaine.edu manomet.org sunrisecounty.org wccog.net Contents 2 Introduction 2 Challenges for River Herring 4 What can be done? 5 What are River Herring? 6 Benefits of River Herring? 7 Managment 9 Status in 2018 11 Restoration Activities in Eastern Maine 14 Case Studies in River Herring Restoration 14 Bagaduce River 17 Patten Stream 17 Union River OUR PARTNERS 18 Narraguagus River 18 East Machias River 21 Orange River 21 Pennamaquan River 22 St. Croix River 23 Looking Ahead DOWNEAST FISHERIES parTNERSHIP / UP AND UP: RIVER HerrING IN EASTERN MAINE 1 Introduction River herring runs in eastern Maine are coming back! A string of successful projects, from the opening of the St. Croix fishway to the renewal of commercial fishing on Card Mill Stream in Franklin, have fueled growing momentum for restoration of rivers and streams, big and small. This report is an update on the status of river herring runs in eastern Maine. It describes the value of river herring and some of the many projects underway to allow river herring to swim upstream to their spawning habitat. Because river herring are a keystone species in freshwater and marine systems—and are so responsive to restoration efforts—they have become a focus of the Downeast Fisheries Partnership effort to restore fisheries in eastern Maine. -

Penobscot River Mercury Study
Case 1:00-cv-00069-JAW Document 699 Filed 12/17/13 Page 1 of 27 PageID #: 4383 Penobscot River Mercury Study Results of 2012 monitoring of mercury in Penobscot River and Bay With comparisons to previous years Report to: Judge John Woodcock U.S. District Court (District of Maine) December 2013 By A.D. Kopec Biologist Penobscot River Mercury Study R.A. Bodaly Project Leader Penobscot River Mercury Study J.W.M. Rudd President, R&K Research Limited, British Columbia, Canada Chair, Penobscot River Mercury Study Panel N.S. Fisher Distinguished Professor, School of Marine and Atmospheric Sciences, Stony Brook University, Stony Brook, NY Member, Penobscot River Mercury Study Panel C.G. Whipple Principal, Environ International Corporation, Emeryville, CA Member, Penobscot River Mercury Study Panel Fish, Shellfish, Water and Sediment Collections K. Payne, C. Francis, M. Bowen, M. Dassatt, and staff Normandeau Associates, Falmouth, ME Bird Collections G. Mittelhauser, A. Leopold, and staff Maine Natural History Observatory, Gouldsboro, ME Black Duck Collections K. Sullivan, B. Allen, H. Cady, and staff Maine Department of Inland Fisheries and Wildlife, Bangor, ME Case 1:00-cv-00069-JAW Document 699 Filed 12/17/13 Page 2 of 27 PageID #: 4384 2 EXECUTIVE SUMMARY The purpose of this report is to present data from the monitoring of mercury (Hg) in water, sediments and biota from 2012, and compare those data to those from previous years in the Penobscot system. Sampling in 2012 represented the second year (after 2010) of the proposed long term monitoring program of Hg in the Penobscot. Results from a number of sampling efforts prior to 2010 were also included where appropriate. -

Recovery Plan for the Gulf of Maine Distinct Population Segment of Atlantic Salmon (Salmo Salar)
Final Recovery Plan for the Gulf of Maine Distinct Population Segment of Atlantic Salmon (Salmo salar) November, 2005 Prepared by NOAA’s National Marine Fisheries Service (NMFS) Silver Spring, Maryland and Northeastern Region U.S. Fish and Wildlife Service Hadley, Massachusetts Recovery Plan for the Gulf of Maine Distinct Population Segment of Atlantic Salmon (Salmo salar) November, 2005 Prepared by NOAA's National Marine Fisheries Service (NMFS) Silver Spring, Maryland and Northeastern Region U.S. Fish and Wildlife Service Hadley, Massachusetts Approved: Assistant Administrator for Fisheries Date NOAA's National Marine Fisheries Service Regional Director, U.S. Fish and Wildlife Service Date DISCLAIMER Recovery plans delineate actions that are thought to be necessary to recover andlor protect endangered species. Recovery plans are prepared by NOAA's National Marine Fisheries Service (NMFS) and the U.S. Fish and Wildlife Service (FWS) and sometimes with the assistance of recovery teams, contractors, state agencies and others. This Recovery Plan for the Gulf of Maine (GOM) Distinct Population Segment (DPS) of Atlantic Salmon (Salmo salar) was prepared by the staff of the Northeast Regional Offices of NMFS with the assistance of the FWS and the Maine Atlantic Salmon Commission (ASC). While the State of Maine provided recommendations for this plan, it was developed using federal guidelines and policies pertaining to recovery plans for federally listed species. Recovery plans are not regulatory or decision documents. The recommendations in a recovery plan are not considered final decisions unless and until they are actually proposed for implementation. Objectives will only be attained and hnds expended contingent upon appropriations, priorities and other budgetary constraints. -

Maine Open Water Fishing Laws Summary, 1975 Maine Department of Inland Fisheries and Game
Maine State Library Digital Maine Inland Fisheries and Wildlife Law Books Inland Fisheries and Wildlife 1-1-1975 Maine Open Water Fishing Laws Summary, 1975 Maine Department of Inland Fisheries and Game Follow this and additional works at: https://digitalmaine.com/ifw_law_books Recommended Citation Maine Department of Inland Fisheries and Game, "Maine Open Water Fishing Laws Summary, 1975" (1975). Inland Fisheries and Wildlife Law Books. 301. https://digitalmaine.com/ifw_law_books/301 This Text is brought to you for free and open access by the Inland Fisheries and Wildlife at Digital Maine. It has been accepted for inclusion in Inland Fisheries and Wildlife Law Books by an authorized administrator of Digital Maine. For more information, please contact [email protected]. Maine Open Water Fishing Laws Summary Maine Department of Inland Fisheries and Game 284 State Street, Augusta, Maine 04330 0££K) auie^M 'e+snSny '+994S 9+p+S VQZ FISHING LICENSES Commissioner of Indian Affairs stating that the person described is an awe0 pue sauaijsij pupguj jo juauupiedaQ auip^Aj Indian and a member of that tribe. For this purpose, an Indian shall RESIDENT NONRESIDENT be any member on tribal lists of the Penobscot and Passamaquoddy 16 years and older . $ 6.50 Season: age 16 and older $15.50 Tribes, or a person on the membership list of the Association of Combination ............. 10.50 12-15 years inclusive .. 2.50 Aroostook Indians and who has resided in Maine for at least 5 years. ‘ 3-day . ’ 6.50 15-day* “ ........................ 10.50 “ Leave or furlough . 2.50 7-day ................................. 7.50 3-day ................................ -

Penobscot Bay Watch
PENOBSCOTBAY Maine National Estuary Program Nomination STATE OF MAINE OFFICEOF THE GOVERNOR AUGUSTA. MAINE 04333 ANGUS S. KING, JR. GOVERNOR March 6, 1995 Administrator Carol Browner US. Environmental Protection Agency 401 M. Street, S.W. Washington, D.C. 20460 Dear Ms. Browner: 1 am pleased to submit a State of Maine Conference Agreement that nominates Penobscot Bay for designation in the National Estuary Program. The Agreement describes the Bay's key issues and actions to respond to them, how we will continue to involve the public in developing and implementing solutions to these most pressing issues, and how we propose to finance this estuary project. Our nomination for Penobscot Bay is consistent with EPA's December; 1994 guidance that calls for considerable initiative by a state before requesting designation. Public and private entities have worked cooperatively since 1992 and made remarkable progress. Two major conferences have brought together all levels of government, the nonprofit community, Bay businesses and the public to talk openly about key issues and possible solutions. These and other activities described in the attached document have laid a solid foundation for a successful national estuaq project. I believe our estuary project will be successfully implemented because of the active participation of five state agencies over the past three years. These agencies (the Departments of Environmental Protection, Marine Resources, Inland Fisheries and Wildlife. Economic and Community Developmenf and the State Planning Office) are committed to this project and I am confident that in concert with our nonprofit, municipal and business partners we will m&e important strides to protect and restore Penobscot Bay. -

Survey of Hancock County, Maine Samuel Wasson
The University of Maine DigitalCommons@UMaine Maine History Documents Special Collections 1878 Survey of Hancock County, Maine Samuel Wasson Follow this and additional works at: https://digitalcommons.library.umaine.edu/mainehistory Part of the United States History Commons Repository Citation Wasson, Samuel, "Survey of Hancock County, Maine" (1878). Maine History Documents. 37. https://digitalcommons.library.umaine.edu/mainehistory/37 This Book is brought to you for free and open access by DigitalCommons@UMaine. It has been accepted for inclusion in Maine History Documents by an authorized administrator of DigitalCommons@UMaine. For more information, please contact [email protected]. SURVEY OF HANCOCK COUNTY. A SURVEY OF HANCOCK COUNTY, MAINE BY SAMIUEL WASSON. MEMBER OF STATE BOARD OK AGRICULTURE. AUGUSTA: SPRAGUE, OWEN A NASH, PRINTERS TO THE STATE. 1878. PREFACE. At the meeting of the Board of Agriculture held at Calais. a resolution was passed, urging the importance to our agri cultural literature of the publication of surveys of the differ ent counties in the State, giving brief notes of their history, industrial resources and agricultural capabilities ; and direct ing the Secretary to procure such contributions for the annual reports. In conformity with this resolution, and also as ear ning out the settled policy of the Board in this respect— evidences of which are found in the publication of similar reports in previous volumes—I give herewith a Survey of the County of Hancock, written by a gentleman who has been a member of the Board of Agriculture, uninterruptedly, from its first organization, and who is in every way well fitted for the work, which he has so well performed. -

Natural Resource Inventory
Natural Resource Inventory Conservation Lands, Sears Island Searsport, Maine For: Friends of Sears Island c/o Marietta Ramsdell PO Box 222, Searsport 04974 Ph. 207 548-0142 [email protected] By: Alison C. Dibble, Ph.D. Jake Maier Stewards LLC JM Forestry P.O. Box 321 6 Lower Falls Road Brooklin, ME 04616 Orland, ME 04472 Ph. 207 359-4659 [email protected] Ph. 207 469-0231 [email protected] http://jmforestry.com 18 January 2011 SUMMARY During the growing season in 2010, Stewards LLC and JM Forestry conducted at Sears Island a natural resource inventory of the new 601-ac conservation easement, held by Maine Coast Heritage Trust. This is located in the lower Penobscot River in Searsport, Waldo County, Maine. As of the 1970s, several large industrial development schemes were proposed for Sears Island. Since the 1990s the island has belonged to the state of Maine. A paved causeway constructed in the 1980s provides year-round access to a gate at the north end of the island. In January 2009 the Friends of Sears Island (FOSI), a nonprofit organization, assumed responsibility for stewardship of the conservation area and this inventory is part of the management planning process. A 70- acre portion of the conservation area includes a vernal pool and a potential building site intended for a future education/ visitor center. The remaining acreage is former farm fields, mature and successional forest, wet forest, streams, and tidal shore. An archaeological site at the north end of the island was documented with an extensive report in 1983. Historic land uses included agriculture and seasonal residences, but no structures remain. -
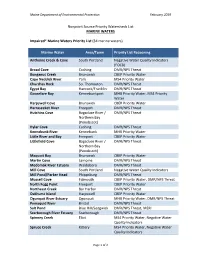
Nonpoint Source Priority Watersheds List MARINE WATERS
Maine Department of Environmental Protection February 2019 Nonpoint Source Priority Watersheds List MARINE WATERS Impaired* Marine Waters Priority List (34 marine waters) Marine Water Area/Town Priority List Reasoning Anthoine Creek & Cove South Portland Negative Water Quality Indicators (FOCB) Broad Cove Cushing DMR/NPS Threat Bunganuc Creek Brunswick CBEP Priority Water Cape Neddick River York MS4 Priority Water Churches Rock So. Thomaston DMR/NPS Threat Egypt Bay Hancock/Franklin DMR/NPS Threat Goosefare Bay Kennebunkport MHB Priority Water, MS4 Priority Water Harpswell Cove Brunswick CBEP Priority Water Harraseeket River Freeport DMR/NPS Threat Hutchins Cove Bagaduce River / DMR/NPS Threat Northern Bay (Penobscot) Hyler Cove Cushing DMR/NPS Threat Kennebunk River Kennebunk MHB Priority Water Little River and Bay Freeport CBEP Priority Water Littlefield Cove Bagaduce River / DMR/NPS Threat Northern Bay (Penobscot) Maquoit Bay Brunswick CBEP Priority Water Martin Cove Lamoine DMR/NPS Threat Medomak River Estuary Waldoboro DMR/NPS Threat Mill Cove South Portland Negative Water Quality Indicators Mill Pond/Parker Head Phippsburg DMR/NPS Threat Mussell Cove Falmouth CBEP Priority Water, DMR/NPS Threat North Fogg Point Freeport CBEP Priority Water Northeast Creek Bar Harbor DMR/NPS Threat Oakhurst Island Harpswell CBEP Priority Water Ogunquit River Estuary Ogunquit MHB Priority Water, DMR/NPS Threat Pemaquid River Bristol DMR/NPS Threat Salt Pond Blue Hill/Sedgwick DMR/NPS Threat, MERI Scarborough River Estuary Scarborough DMR/NPS Threat Spinney Creek Eliot MS4 Priority Water, Negative Water Quality Indicators Spruce Creek Kittery MS4 Priority Water, Negative Water Quality Indicators Page 1 of 2 MDEP NPS Priority Watersheds List – MARINE WATERS February 2019 Marine Water Area/Town Priority List Reasoning Spurwink River Scarborough MHB Priority Water, DMR/NPS Threat St. -

A History of Oysters in Maine (1600S-1970S) Randy Lackovic University of Maine, [email protected]
The University of Maine DigitalCommons@UMaine Darling Marine Center Historical Documents Darling Marine Center Historical Collections 3-2019 A History of Oysters in Maine (1600s-1970s) Randy Lackovic University of Maine, [email protected] Follow this and additional works at: https://digitalcommons.library.umaine.edu/dmc_documents Part of the Aquaculture and Fisheries Commons, History of Science, Technology, and Medicine Commons, and the United States History Commons Repository Citation Lackovic, Randy, "A History of Oysters in Maine (1600s-1970s)" (2019). Darling Marine Center Historical Documents. 22. https://digitalcommons.library.umaine.edu/dmc_documents/22 This Newsletter is brought to you for free and open access by DigitalCommons@UMaine. It has been accepted for inclusion in Darling Marine Center Historical Documents by an authorized administrator of DigitalCommons@UMaine. For more information, please contact [email protected]. A History of Oysters in Maine (1600s-1970s) This is a history of oyster abundance in Maine, and the subsequent decline of oyster abundance. It is a history of oystering, oyster fisheries, and oyster commerce in Maine. It is a history of the transplanting of oysters to Maine, and experiments with oysters in Maine, and of oyster culture in Maine. This history takes place from the 1600s to the 1970s. 17th Century {}{}{}{} In early days, oysters were to be found in lavish abundance along all the Atlantic coast, though Ingersoll says it was at least a small number of oysters on the Gulf of Maine coast.86, 87 Champlain wrote that in 1604, "All the harbors, bays, and coasts from Chouacoet (Saco) are filled with every variety of fish.