Sanctuary Science Report 2001: an Ecosystem Report Card
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Highlights Report with Live Links
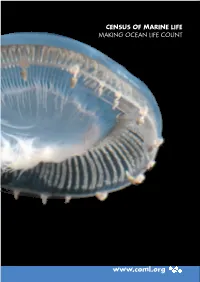
CENSUS OF MARINE LIFE MAKING OCEAN LIFE COUNT www.coml.org Census of Marine Life Projects Information System: OBIS Nearshore: NaGISA Coral Reefs: CReefs Regional Ecosystems: GoMA Continental Shelves: POST Continental Margins: COMARGE Abyssal Plains: CeDAMar Mid-Ocean Ridges: MAR-ECO Seamounts: CenSeam Vents and Seeps: ChEss Arctic Ocean: ArcOD Antarctic Ocean: CAML Top Predators: TOPP Zooplankton: CMarZ Microbes: ICoMM Oceans Past: HMAP Oceans Future: FMAP National and Regional Implementation Committees Australia Canada Caribbean China Europe Japan Indian Ocean Indonesia South Korea South America Sub-Saharan Africa United States Like an underwater spaceship, a jellyfish,Aequorea macrodactyla, travels through the warm, clear waters of the Celebes Sea in the Western Pacific Ocean. The jellyfish was but one of thousands of specimens photographed during a three-week Census expedition to explore this highly diverse area. Photo: Larry Madin, Woods Hole Oceanographic Institution. A Truly Global Endeavor The Census of Marine Life is a global network of researchers in more than 80 nations engaged in a ten-year scientific initiative to assess and ex- plain the diversity, distribution, and abundance of marine life in the oceans. The world’s first com- prehensive Census of Marine Life—past, present, and future—will be released in 2010. The Census gratefully acknowledges the financial support of numerous governments and organiza- tions from around the world. Moreover, many of the highlights in this report were only realized through the generous collaborative spirit and un- precedented cooperation of Census researchers and their international colleagues. A complete list of Census sponsors, funding partners, collaborat- ing institutions, and participating individuals is available at www.coml.org. -

Guide to Theecological Systemsof Puerto Rico
United States Department of Agriculture Guide to the Forest Service Ecological Systems International Institute of Tropical Forestry of Puerto Rico General Technical Report IITF-GTR-35 June 2009 Gary L. Miller and Ariel E. Lugo The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the National Forests and national grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable sex, marital status, familial status, parental status, religion, sexual orientation genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD).To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, 1400 Independence Avenue, S.W. Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. Authors Gary L. Miller is a professor, University of North Carolina, Environmental Studies, One University Heights, Asheville, NC 28804-3299. -

AMNH Digital Library
lime • Fish tha 10/04 V / / / X Come Fly $3.95 USA $4.95 CANADA 1 0> WITH Me! 246846 4 M*"-^- ^'.'.'L-^ -m We're not just breaking new ground W I ! I I ^J U I V W III W I \^ W Toyota's eight U.S. manufacturing plants are setting the standards for building quality into every vehicle we produce. Today, eight Toyota models are manufactured in the U.S. and last year alone we built more than a million vehicles here.* Since we first began doing business in America, Toyota has been committed to reinvesting into the local economy. Through manufacturing, sales and marketing, research and design facilities, as well as our purchasing from local suppliers, we're creating jobs and opportunities in communities across America. And with new plants under construction in San Antonio, Texas and Jackson, Tennessee, we'll continue to break new ground every day. "Toyota components and vehicles are made using many U.S. sourced parts. ^§'2004 toyota.com/usa TOYOTA ALAPAGOS CXpGiriGIICG IVIdttGirSa And so does commitment. I first visited Galapagos with my fattier in 1967, when he opened up the possibility of exploring the islands by ship. I still remember that voyage moment by moment. I was struck above all with the wildlife's total lack of fear as my daughter (below) discovered too in her first encounter with marine iguanas. Galapagos is a very special place and I believe that if you, the traveler, have a great experience there, your passion will play a big role in securing the future well-being of these islands. -

Host Specificity and Phylogenetic Relationships Among Atlantic Ovulidae (Mollusca: Gastropoda)
Contributions to Zoology, 79 (2) 69-78 (2010) Host specificity and phylogenetic relationships among Atlantic Ovulidae (Mollusca: Gastropoda) Bastian T. Reijnen1, 2, Bert W. Hoeksema1, Edmund Gittenberger1 1 Department of Marine Zoology, Netherlands Centre for Biodiversity Naturalis, PO Box 9517, NL 2300 RA Leiden, The Netherlands 2 E-mail: [email protected] Key words: Anthozoa, Alcyonacea, Caribbean, Curaçao, molecular phylogeny, symbiosis Abstract the Ovulidae for a long time (Goud and Hoeksema, 2001), live on stylasterid corals (Hydrozoa: Athecatae: Ovulid gastropods and their octocoral hosts were collected Filefera). Pedicularia differs from the undisputed along the leeward coast of Curaçao, Netherlands Antilles. New molecular data of Caribbean and a single Atlantic species were ovulids in radula morphology (Simone, 2004); it is combined with comparable data of Indo-Pacific Ovulidae and a now classified in the separate family Pediculariidae single East-Pacific species from GenBank. Based on two DNA (Fehse, 2007; Lorenz and Fehse, 2009). markers, viz. CO-I and 16S, the phylogenetic relationships Thirty-seven species of Ovulidae Fleming, 1822, are among all ovulid species of which these data are available are known from the Caribbean and Atlantic area (Lorenz reconstructed. The provisional results suggest a dichotomy be- and Fehse, 2009). The dominant genus in the Carib- tween the Atlantic and the Indo-Pacific taxa. Fully grownSimnia - lena uniplicata closely resembles juvenile Cyphoma gibbosum bean is Cyphoma Röding, 1798, with 14 species of conchologically. Cymbovula acicularis and C. bahamaensis which Cyphoma gibbosum (Linnaeus, 1758) is the might be synonyms. The assignments of Caribbean host species most common. Due to the low number of ovulid spe- for Cyphoma gibbosum, C. -

Mesopredator Release and the Effectiveness of Camera Traps for Estimating Mammal Abundances in the Eastern Cape, South Africa
Mesopredator release and the effectiveness of camera traps for estimating mammal abundances in the Eastern Cape, South Africa A thesis submitted in fulfilment of the requirements for the degree of MASTER OF SCIENCE of RHODES UNIVERSITY By JONATHAN MICHAEL TAYLOR November 2015 Abstract Remotely operated cameras (camera traps) have become an indispensable tool for many ecologists, particularly those studying rare and elusive animals. A plethora of camera trap makes and models are now commercially available, yet the effects of their varying design features on the quality and quantity of data recorded remains principally unknown. Better understanding of differing camera trap designs is needed before adequate management policies can be implemented, especially when the aim is to protect vulnerable and endangered species such as many carnivores. Habitat loss and human conflict has prompted worldwide declines of apex predator populations. Following this, many smaller predators have undergone population ‘explosions’ due to the lack of top-down forcing, a phenomenon known as mesopredator release. Land use changes in the Eastern Cape of South Africa have caused extensive degradation and fragmentation of the Subtropical Thicket Biome. In addition, coupled with anthropogenic persecution, apex predators including lions ( Panthera leo ), leopards ( P. pardus ), and brown hyaenas ( Hyaena brunnea ) have been extirpated from large areas of their historic range. Removal of these apex predators may provide opportunity for mesopredators, such as black-backed jackals ( Canis mesomelas ) and caracals ( Caracal caracal ), to be released from top-down forcing and possibly initiate trophic cascade effects. Therefore, using randomly distributed camera traps, my study aimed to explore the quality and quantity of data collected by different camera trap designs, and to investigate the mesopredator release hypothesis in the Eastern Cape of South Africa. -

Isotopic Labeling and Antifungal Resistance As Tracers of Gut Passage of the Sea Fan Pathogen Aspergillus Sydowii
Vol. 86: 1–7, 2009 DISEASES OF AQUATIC ORGANISMS Published September 7 doi: 10.3354/dao02106 Dis Aquat Org Isotopic labeling and antifungal resistance as tracers of gut passage of the sea fan pathogen Aspergillus sydowii Krystal L. Rypien1, 2,*, David M. Baker1 1Department of Ecology and Evolutionary Biology, Dale R. Corson Hall, Cornell University, Ithaca, New York 14853, USA 2Present address: Scripps Institution of Oceanography, Marine Biology Research Division, 9500 Gilman Drive MC 0202, La Jolla, California 92093, USA ABSTRACT: Vectors play a critical role in the ecology of infectious disease by facilitating between- host transmission, emphasizing the multi-species nature of disease. Corals are suffering an onslaught of infectious diseases, yet we know little about the role of vector species in the ecology of these epi- zootics. The infection of octocorals by the fungus Aspergillus sydowii is a widespread Caribbean coral disease. The snail Cyphoma gibbosum is a likely vector species because it is a specialist preda- tor of octocorals, moves regularly among coral colonies, and aggregates on diseased corals. We used a novel application of stable isotope techniques and antifungal-resistant strains of A. sydowii to iden- tify the potential for this snail to vector disease. The use of both isotopically labeled and hygromycin- resistant fungus was successful in definitively tracing A. sydowii through the guts of C. gibbosum from ingestion to feces. Further, we demonstrated in laboratory experiments that snails fed antifun- gal-resistant A. sydowii excrete viable spores and hyphae in their feces. Overall, this study demon- strates the usefulness of isotopic labeling and antifungal-resistant fungi to trace the movement of pathogen propagules in microbially diverse environments, and suggests that C. -

ISSUE 4 - 2017 Newsbio Editor’S Letter Dutch Caribbean, October 2017 BIONEWS ISSUE 4
BIONEwS ISSUE 4 - 2017 Editor’s Letter Dutch Caribbean, October 2017 BIONEWS ISSUE 4 2 Editor’s Letter As part of DCNA’s series of articles focusing on the The scientific journalMarine Biodiversity has status of reefs in all five of our islands, we here put recently focused its attention on the reefs of the 3 Status of Curaçao’s Reefs the spotlight on Curaçao. The island’s reefs have Dutch Caribbean with various articles reporting been the subject of much research in the past few on recent coral reef research in St. Eustatius and 12 Researchers discover that the years, and in this edition of BioNews we attempt Curaçao, such as a report on the deep reef three known flamingo tongue snail to provide insight into some of the new findings. community (70-85 m depth) discovered off the species are in fact just one species Commonly described as healthy and diverse and leeward coast of Curaçao. We highly encourage praised by divers and snorkelers for their beauty you to look at the list with all articles published by 13 Biodiversity of and rich marine fauna, the island’s reefs appear to Marine Biodiversity on the Dutch Caribbean to find Dutch Caribbean Reefs be following the Caribbean-wide trend of declin- out more about the many new discoveries that ing reef health. We look at the findings of several have been made! 14 Research Overview recent studies that highlight the significant shift that the island’s coral reef communities have gone One exciting discovery was recently made by two 16 Long-Term Projects Overview through over the past four decades, with a notable researchers from Naturalis and Oxford University loss in both coral cover and fish biomass and a wor- Museum of Natural History about the flamingo 24 Monitoring Overview rying increase in turf algae. -

Bionews ISSUE 4 - 2017
BIONEwS ISSUE 4 - 2017 In this issue: 1 Editor’s Letter 16 Long-Term Projects Overview 2 Status of Curaçao’s Reefs 24 Monitoring Overview 11 Researchers discover that the three 28 List of Acronyms known flamingo tongue snail species are in fact just one species 29 Reports and Publications 12 Biodiversity of 30 Calendar Dutch Caribbean Reefs 31 Members and Contact 13 Research Overview 32 References Editor’s Letter As part of DCNA’s series of articles focusing on as pollution and coastal development have had the status of reefs in all five of our islands, we a drastic impact on reef health.The scientific here put the spotlight on Curaçao. The island’s journal Marine Biodiversity has recently focused reefs have been the subject of much research its attention on the reefs of the Dutch Caribbean in the past few years, and in this edition of with various articles reporting on recent coral reef BioNews we attempt to provide insight into research in St. Eustatius and Curaçao, such as a some of the new findings. Commonly described report on the deep reef community (70-85 m as healthy and diverse and praised by divers depth) discovered off the leeward coast of and snorkelers for their beauty and rich marine Curaçao. We highly encourage you to look at fauna, the island’s reefs appear to be follow- the list with all articles published by Marine ing the Caribbean-wide trend of declining reef Biodiversity on the Dutch Caribbean to find out health. We look at the findings of several recent more about the many new discoveries that have studies that highlight the significant shift that been made! the island’s coral reef communities have gone through over the past four decades, with a One exciting discovery was recently made by two notable loss in both coral cover and fish biomass researchers from Naturalis and Oxford University and a worrying increase in turf algae. -
European Coral Reef Symposium Oxford, 2017
European Coral Reef Symposium Oxford, 2017 V.2. 1 Contents Welcome Dear Delegates, 3. Welcome Welcome to the European Coral Reef Symposium! 4. Brought to you by We, the Reef Conservaton UK (RCUK) commitee, were invited by the Internatonal Society of Reef Studies to host a European coral reef meetng in 2017 in celebraton of twenty years of RCUK. Almost a 5. ECRS organising commitee year later and over 500 coral reef nerds are descending on a very festve Oxford! 6. ECRS Volunteers The interest in the conference has been overwhelming, it has pulled us out of our traditonal (yet 7. Sustainable conference smaller) home at ZSL London Zoo and into something rather more prestgious. The friendly and relaxed atmosphere of our annual gatherings is at the heart of RCUK and we hope this has travelled with us to 8. The Venues Oxford. 9. Moving between venues We have strived to make ECRS2017 as sustainable, environmentally friendly and family friendly as possible. Please join us on the band wagon by bringing along your refllable water botle and downloading 10. Inside the Examinaton Schools the conference App. 11. Conference venue map We look forward to seeing you all and hope you enjoy three days of sharing science! 12. Our plenary speakers The ECRS/RCUK commitee 13. Informaton for presenters #ECRS2017 [email protected] 14 - 17. The Schedule @ReefConsUK www.reefconservatonuk.co.uk/ecrs 18 - 37. Session line-ups 38 - 40. Workshop informaton 20 years of Reef Conservaton UK (RCUK) 41. Student grants & prizes Founding RCUK commitee Kristan Teleki, Heather Koldewey, Alastair Harborne, Dorie Smith, Jan Smith, Damon Stanwell-Smith, Elizabeth 42 - 45. -

Charlotteville Bioblitz 2015 Final Report.Pdf
Final Report Contents Report Credits ....................................................................................................... iii Executive Summary ................................................................................................ 1 Introduction ........................................................................................................... 2 Methods Plants......................................................................................................... 3 Birds .......................................................................................................... 3 Mammals .................................................................................................. 4 Reptiles and Amphibians .......................................................................... 4 Aquatic ...................................................................................................... 4 Terrestrial Invertebrates ........................................................................... 5 Fungi .......................................................................................................... 7 Results and Discussion Plants......................................................................................................... 7 Birds .......................................................................................................... 8 Mammals .................................................................................................. 9 Reptiles and Amphibians ......................................................................... -

Part 2 – Coral Reef Ecology
PART 2 – CORAL REEF ECOLOGY Coral reefs are rocky mounds and/or ridges formed in the sea by marine organisms through the accumulation and deposition of limestone (calcium carbonate). The reef framework provides the structural foundation of a unique and rich marine ecosystem. A single reef may cover over100 sq km. The living reef forms the top layer of the reef adding new limestone to the reef. These specialized habitats provide shelter, food, and breeding sites for numerous plants and animals and form a breakwater for the adjacent coast, providing natural storm protection Great Barrier Reef Stony corals are the major reef architects. These small marine animals, (individual organisms are called polyps), produce a hard skeleton made of calcium carbonate, which they extract from the seawater and combine with CO2 for limestone. Coral Types Hard (Stony, scleractinian, “true”) corals build the reef by extracting calcium carbonate from the ocean water and they create a diverse 3-D habitat for many other organisms Coral are actually a special group of cnidarians Hermatypic Corals - Corals that form large colonies called reefs and have a symbiotic relationship with the dinoflagellate Zooxanthellae Ahermatypic Corals - Corals that are solitary or form small colonies- they often lack the symbiotic relationship with Zooxanthellae and do not help build reefs All the different colors and shapes made up of thousands of individual polyps, each secreting its own small cup of coral limestone, which provide the building blocks for reef construction. -
Ciclo Reproductivo Del Caracol Pala Strombus Gigas Linnaeus 1758
CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS DEL INSTITUTO POLITÉCNICO NACIONAL CINVESTAV-IPN UNIDAD MÉRIDA Departamento de Recursos del Mar Ciclo reproductivo del Caracol pala Strombus gigas Linnaeus 1758 (Gastropoda: Caenogastropoda: Strombidae) del Archipiélago de San Andrés, Providencia y Santa Catalina, Caribe insular colombiano Tesis que presenta Omar Hernando Avila Poveda Para obtener el grado de Maestro en Ciencias En la especialidad de Biología Marina Directora de Tesis: Dra. Luz Maria Dalila Aldana Aranda Mérida, Yucatán septiembre, 2004 Ciclo reproductivo de Strombus gigas del Caribe insular colombiano © Derechos de autor 2004, OMAR HERNANDO AVILA-POVEDA [email protected] Bogotá, Colombia. A mi amada esposa y futuro retoñito(a) A mis padres, hermanos, sobrino y ahijado i Omar Hernando Avila Poveda RESUMEN La falta de estudios directos sobre el ciclo reproductivo del Caracol pala Strombus gigas, han dificultado plantear medidas de regulación acordes a su ciclo; que permitan el uso racional y sustentable de este recurso comunitario para todo el Caribe. Esta tesis investiga por medio del análisis histológico de la gónada el ciclo reproductivo y la época de desove de Strombus gigas de los cayos del sur del Archipiélago de San Andrés, Providencia y Santa Catalina, Caribe insular colombiano. Adicional a este estudio, se presenta: la proporción de sexos, la longitud de madurez (Lma t), el grosor de madurez (Gmat), y la relación grosor del labio-longitud de la concha mediante un modelo lineal simple con datos transformados a logaritmo natural (Ln[G] = a + bLn[L]). Fueron analizados 346 organismos colectados desde febrero de 2003 hasta enero de 2004. Se tomaron datos biométricos (longitud total de la concha y grosor del labio de la concha) y muestras de masa visceral (gónada y glándula digestiva).