Open Full Page
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
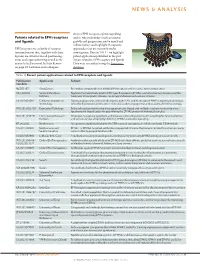
Patents Related to EPH Receptors and Ligands
NEWS & ANALYSIS discuss EPH receptor–ephrin signalling Patents related to EPH receptors and its role in disorders such as tumour and ligands growth and progression, nerve injury and inflammation, and highlight therapeutic EPH receptors are a family of receptor approaches that are currently under tyrosine kinases that, together with their investigation. Here in TABLE 1 we highlight ligands, are involved in cell positioning, patent applications published in the past tissue and organ patterning as well as the 3 years related to EPH receptors and ligands. control of cell survival. In their Review Data were researched using the Espacenet on page 39, Lackman and colleagues database. Table 1 | Recent patent applications related to EPH receptors and ligands Nature Reviews | Drug Discovery Publication Applicants Subject numbers NZ 581397 AstraZeneca Pyrimidine compounds that inhibit EPH receptors and are useful for treating cancer HK 1108702 Sanford-Burnham Peptides that selectively bind to EPH type-B receptors (EPHBs); useful for tumour imaging and the Institute treatment of neoplastic disease, neurological disease and vascular disease US 2013091591 California Institute of During angiogenesis, arterial cells express ephrin B2, and its receptor EPHB4 is expressed on venous Technology cells; this distinction can be used in methods to alter angiogenesis and to assess the effect of drugs WO 2013052710 Expression Pathology Selected reaction monitoring mass spectrometry-based and multiple reaction monitoring mass spectrometry-based assays for quantifying -

A Review of VEGF/VEGFR-Targeted Therapeutics for Recurrent Glioblastoma
414 Original Article A Review of VEGF/VEGFR-Targeted Therapeutics for Recurrent Glioblastoma David A. Reardon, MDa,b; Scott Turner, MDc; Katherine B. Peters, MD, PhDc; Annick Desjardins, MDc; Sridharan Gururangan, MDa,b; John H. Sampson, MD, PhDa; Roger E. McLendon, MDd; James E. Herndon II, PhDe; Lee W. Jones, PhDf; John P. Kirkpatrick, MD, PhDf; Allan H. Friedman, MDa; James J. Vredenburgh, MDc; Darell D. Bigner, MD, PhDd; and Henry S. Friedman, MDa,b; Durham, North Carolina Key Words ability effect of VEGF inhibitors, the Radiologic Assessment in Neu- Glioblastoma, angiogenesis, vascular endothelial growth factor, ro-Oncology (RANO) criteria were recently implemented to bet- malignant glioma ter assess response among patients with glioblastoma. Although bevacizumab improves survival and quality of life, eventual tumor progression is the norm. Better understanding of resistance mech- Abstract anisms to VEGF inhibitors and identification of effective therapy Glioblastoma, the most common primary malignant brain tumor after bevacizumab progression are currently a critical need for pa- among adults, is a highly angiogenic and deadly tumor. Angiogen- tients with glioblastoma. (JNCCN 2011;9:414–427) esis in glioblastoma, driven by hypoxia-dependent and indepen- dent mechanisms, is primarily mediated by vascular endothelial growth factor (VEGF), and generates blood vessels with distinctive features. The outcome for patients with recurrent glioblastoma is poor because of ineffective therapies. However, recent encourag- ing rates of radiographic response and progression-free survival, Malignant gliomas, including the most common sub- and adequate safety, led the FDA to grant accelerated approval of type of glioblastoma, are rapidly growing destructive tu- bevacizumab, a humanized monoclonal antibody against VEGF, for mors that extensively invade locally but rarely metasta- the treatment of recurrent glioblastoma in May 2009. -

A Tyrosine Kinase Expression Signature Predicts the Post-Operative Clinical Outcome in Triple Negative Breast Cancers
cancers Article A Tyrosine Kinase Expression Signature Predicts the Post-Operative Clinical Outcome in Triple Negative Breast Cancers Alexandre de Nonneville 1, Pascal Finetti 2 , José Adelaide 2, Éric Lambaudie 3, Patrice Viens 1, Anthony Gonçalves 1 , Daniel Birnbaum 2, Emilie Mamessier 2 and François Bertucci 1,2,* 1 Department of Medical Oncology, Institut Paoli-Calmettes, Aix-Marseille Univ, CRCM, CNRS, INSERM, 13000 Marseille, France 2 Laboratory of Predictive Oncology, Centre de Recherche en Cancérologie de Marseille, Institut Paoli-Calmettes, Inserm UMR1068, CNRS UMR725, Aix-Marseille Université, 13000 Marseille, France 3 Department of Surgical Oncology, Institut Paoli-Calmettes, Aix-Marseille Univ, CNRS, INSERM, CRCM, 13000 Marseille, France * Correspondence: [email protected]; Tel.: +33-4-91-22-35-37; Fax: +33-4-91-22-36-70 Received: 15 July 2019; Accepted: 9 August 2019; Published: 13 August 2019 Abstract: Triple negative breast cancer (TNBC) represent 15% of breast cancers. Histoclinical features and marketed prognostic gene expression signatures (GES) failed to identify good- and poor-prognosis patients. Tyrosine kinases (TK) represent potential prognostic and/or therapeutic targets for TNBC. We sought to define a prognostic TK GES in a large series of TNBC. mRNA expression and histoclinical data of 6379 early BCs were collected from 16 datasets. We searched for a TK-based GES associated with disease-free survival (DFS) and tested its robustness in an independent validation set. A total of 1226 samples were TNBC. In the learning set of samples (N = 825), we identified a 13-TK GES associated with DFS. This GES was associated with cell proliferation and immune response. -

Functional Analysis of Somatic Mutations Affecting Receptor Tyrosine Kinase Family in Metastatic Colorectal Cancer
Author Manuscript Published OnlineFirst on March 29, 2019; DOI: 10.1158/1535-7163.MCT-18-0582 Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Functional analysis of somatic mutations affecting receptor tyrosine kinase family in metastatic colorectal cancer Leslie Duplaquet1, Martin Figeac2, Frédéric Leprêtre2, Charline Frandemiche3,4, Céline Villenet2, Shéhérazade Sebda2, Nasrin Sarafan-Vasseur5, Mélanie Bénozène1, Audrey Vinchent1, Gautier Goormachtigh1, Laurence Wicquart6, Nathalie Rousseau3, Ludivine Beaussire5, Stéphanie Truant7, Pierre Michel8, Jean-Christophe Sabourin9, Françoise Galateau-Sallé10, Marie-Christine Copin1,6, Gérard Zalcman11, Yvan De Launoit1, Véronique Fafeur1 and David Tulasne1 1 Univ. Lille, CNRS, Institut Pasteur de Lille, UMR 8161 - M3T – Mechanisms of Tumorigenesis and Target Therapies, F-59000 Lille, France. 2 Univ. Lille, Plateau de génomique fonctionnelle et structurale, CHU Lille, F-59000 Lille, France 3 TCBN - Tumorothèque Caen Basse-Normandie, F-14000 Caen, France. 4 Réseau Régional de Cancérologie – OncoBasseNormandie – F14000 Caen – France. 5 Normandie Univ, UNIROUEN, Inserm U1245, IRON group, Rouen University Hospital, Normandy Centre for Genomic and Personalized Medicine, F-76000 Rouen, France. 6 Tumorothèque du C2RC de Lille, F-59037 Lille, France. 7 Department of Digestive Surgery and Transplantation, CHU Lille, Univ Lille, 2 Avenue Oscar Lambret, 59037, Lille Cedex, France. 8 Department of hepato-gastroenterology, Rouen University Hospital, Normandie Univ, UNIROUEN, Inserm U1245, IRON group, F-76000 Rouen, France. 9 Department of Pathology, Normandy University, INSERM 1245, Rouen University Hospital, F 76 000 Rouen, France. 10 Department of Pathology, MESOPATH-MESOBANK, Centre León Bérard, Lyon, France. 11 Thoracic Oncology Department, CIC1425/CLIP2 Paris-Nord, Hôpital Bichat-Claude Bernard, Paris, France. -

Ephrin-A Binding and Epha Receptor Expression Delineate the Matrix Compartment of the Striatum
The Journal of Neuroscience, June 15, 1999, 19(12):4962–4971 Ephrin-A Binding and EphA Receptor Expression Delineate the Matrix Compartment of the Striatum L. Scott Janis, Robert M. Cassidy, and Lawrence F. Kromer Department of Cell Biology and Interdisciplinary Program in Neuroscience, Georgetown University Medical Center, Washington, DC 20007 The striatum integrates limbic and neocortical inputs to regulate matrix neurons. In situ hybridization for EphA RTKs reveals that sensorimotor and psychomotor behaviors. This function is de- the two different ligand binding patterns strictly match pendent on the segregation of striatal projection neurons into the mRNA expression patterns of EphA4 and EphA7. anatomical and functional components, such as the striosome Ligand–receptor binding assays indicate that ephrin-A1 and and matrix compartments. In the present study the association ephrin-A4 selectively bind EphA4 but not EphA7 in the lysates of ephrin-A cell surface ligands and EphA receptor tyrosine of striatal tissue. Conversely, ephrin-A2, ephrin-A3, and kinases (RTKs) with the organization of these compartments ephrin-A5 bind EphA7 but not EphA4. These observations im- was determined in postnatal rats. Ephrin-A1 and ephrin-A4 plicate selective interactions between ephrin-A molecules and selectively bind to EphA receptors on neurons restricted to the EphA RTKs as potential mechanisms for regulating the com- matrix compartment. Binding is absent from the striosomes, partmental organization of the striatum. which were identified by m-opioid -

Acquired Resistance to Dasatinib in Lung Cancer Cell Lines Conferred by DDR2 Gatekeeper Mutation and NF1 Loss
Published OnlineFirst December 2, 2013; DOI: 10.1158/1535-7163.MCT-13-0817 Molecular Cancer Cancer Biology and Signal Transduction Therapeutics Acquired Resistance to Dasatinib in Lung Cancer Cell Lines Conferred by DDR2 Gatekeeper Mutation and NF1 Loss Ellen M. Beauchamp1, Brittany A. Woods1,7, Austin M. Dulak1, Li Tan3, Chunxiao Xu1, Nathanael S. Gray2, Adam J. Bass1,6, Kwok-kin Wong1,4, Matthew Meyerson1,5,6, and Peter S. Hammerman1,6 Abstract The treatment of non–small cell lung cancer has evolved dramatically over the past decade with the adoption of widespread use of effective targeted therapies in patients with distinct molecular alterations. In lung squamous cell carcinoma (lung SqCC), recent studies have suggested that DDR2 mutations are a biomarker for therapeutic response to dasatinib and clinical trials are underway testing this hypothesis. Although targeted therapeutics are typically quite effective as initial therapy for patients with lung cancer, nearly all patients develop resistance with long-term exposure to targeted drugs. Here, we use DDR2-dependent lung cancer cell lines to model acquired resistance to dasatinib therapy. We perform targeted exome sequencing to identify two distinct mechanisms of acquired resistance: acquisition of the T654I gatekeeper mutation in DDR2 and loss of NF1. We show that NF1 loss activates a bypass pathway, which confers ERK dependency downstream of RAS activation. These results indicate that acquired resistance to dasatinib can occur via both second-site mutations in DDR2 and by activation of bypass pathways. These data may help to anticipate mechanisms of resistance that may be identified in upcoming clinical trials of anti-DDR2 therapy in lung cancer and suggest strategies to overcome resistance. -

Global DNA Methylation Profiling Reveals Silencing of a Secreted Form
Oncogene (2007) 26, 4243–4252 & 2007 Nature Publishing Group All rights reserved 0950-9232/07 $30.00 www.nature.com/onc ORIGINAL ARTICLE Global DNA methylation profiling reveals silencing of a secreted form of Epha7in mouse and human germinal center B-cell lymphomas DW Dawson1, JS Hong1, RR Shen1, SW French1, JJ Troke1, Y-Z Wu2, S-S Chen2, D Gui1, M Regelson3, Y Marahrens3, HC Morse III4, J Said1, C Plass2 and MA Teitell1,5 1Department of Pathology and Laboratory Medicine, David Geffen School of Medicine at UCLA, Los Angeles, CA, USA; 2Molecular Virology, Immunology and Medical Genetics, Division of Human Cancer Genetics, The Ohio State University, Columbus, OH, USA; 3Department of Human Genetics, The David Geffen School of Medicine at UCLA, Los Angeles, CA, USA; 4Laboratory of Immunopathology, National Institute of Allergy and Infectious Disease, National Institutes of Health, Rockville, MD, USA and 5Molecular Biology Institute, Institute for Stem Cell Biology and Medicine, California NanoSystems Institute, Institute for Cell Mimetic Studies and Jonsson Comprehensive Cancer Center, UCLA, Los Angeles, CA, USA Most human lymphomas originate from transformed Introduction germinal center (GC) B lymphocytes. While activating mutations and translocations of MYC, BCL2 and BCL6 The TCL1 oncoprotein augments AKT activation and promote specific GC lymphoma subtypes, other genetic downstream signaling to promote cell proliferation and and epigenetic modifications that contribute to malignant survival (Teitell, 2005). Dysregulated expression of progression in the GC remain poorly defined. Recently, TCL1 is a frequent molecular alteration in non-Hodgkin aberrant expression of the TCL1 proto-oncogene was lymphomas (NHLs) that arise by transformation of identified in major GC lymphoma subtypes. -

EPH/Ephrin Profile and EPHB2 Expression Predicts Patient Survival in Breast Cancer
www.impactjournals.com/oncotarget/ Oncotarget, Vol. 7, No. 16 EPH/ephrin profile and EPHB2 expression predicts patient survival in breast cancer Anna-Maria Husa1,2, Željana Magić1, Malin Larsson3, Tommy Fornander4, Gizeh Pérez-Tenorio1 1Department of Clinical and Experimental Medicine, Division of Oncology, Linköping University, Linköping, Sweden 2Current address: CCRI, Children’s Cancer Research Institute, St. Anna Kinderkrebsforschung e.V., Vienna, Austria 3Bioinformatics Infrastructure for Life Sciences (BILS) and Department of Physics, Chemistry and Biology, Linköping University, Linköping, Sweden 4Department of Oncology, Karolinska University Hospital and Karolinska Institute, Stockholm, Sweden Correspondence to: Gizeh Pérez-Tenorio, e-mail: [email protected] Keywords: EPHB2, EPH family, TaqMan array, gene expression, protein expression Received: July 03, 2015 Accepted: January 23, 2016 Published: February 8, 2016 ABSTRACT The EPH and ephrins function as both receptor and ligands and the output on their complex signaling is currently investigated in cancer. Previous work shows that some EPH family members have clinical value in breast cancer, suggesting that this family could be a source of novel clinical targets. Here we quantified the mRNA expression levels of EPH receptors and their ligands, ephrins, in 65 node positive breast cancer samples by RT-PCR with TaqMan® Micro Fluidics Cards Microarray. Upon hierarchical clustering of the mRNA expression levels, we identified a subgroup of patients with high expression, and poor clinical outcome. EPHA2, EPHA4, EFNB1, EFNB2, EPHB2 and EPHB6 were significantly correlated with the cluster groups and particularly EPHB2 was an independent prognostic factor in multivariate analysis and in four public databases. The EPHB2 protein expression was also analyzed by immunohistochemistry in paraffin embedded material (cohort 2). -

Protein Tyrosine Kinases: Their Roles and Their Targeting in Leukemia
cancers Review Protein Tyrosine Kinases: Their Roles and Their Targeting in Leukemia Kalpana K. Bhanumathy 1,*, Amrutha Balagopal 1, Frederick S. Vizeacoumar 2 , Franco J. Vizeacoumar 1,3, Andrew Freywald 2 and Vincenzo Giambra 4,* 1 Division of Oncology, College of Medicine, University of Saskatchewan, Saskatoon, SK S7N 5E5, Canada; [email protected] (A.B.); [email protected] (F.J.V.) 2 Department of Pathology and Laboratory Medicine, College of Medicine, University of Saskatchewan, Saskatoon, SK S7N 5E5, Canada; [email protected] (F.S.V.); [email protected] (A.F.) 3 Cancer Research Department, Saskatchewan Cancer Agency, 107 Wiggins Road, Saskatoon, SK S7N 5E5, Canada 4 Institute for Stem Cell Biology, Regenerative Medicine and Innovative Therapies (ISBReMIT), Fondazione IRCCS Casa Sollievo della Sofferenza, 71013 San Giovanni Rotondo, FG, Italy * Correspondence: [email protected] (K.K.B.); [email protected] (V.G.); Tel.: +1-(306)-716-7456 (K.K.B.); +39-0882-416574 (V.G.) Simple Summary: Protein phosphorylation is a key regulatory mechanism that controls a wide variety of cellular responses. This process is catalysed by the members of the protein kinase su- perfamily that are classified into two main families based on their ability to phosphorylate either tyrosine or serine and threonine residues in their substrates. Massive research efforts have been invested in dissecting the functions of tyrosine kinases, revealing their importance in the initiation and progression of human malignancies. Based on these investigations, numerous tyrosine kinase inhibitors have been included in clinical protocols and proved to be effective in targeted therapies for various haematological malignancies. -

ALL1 Fusion Proteins Induce Deregulation of Epha7 and ERK Phosphorylation in Human Acute Leukemias
ALL1 fusion proteins induce deregulation of EphA7 and ERK phosphorylation in human acute leukemias Hiroshi Nakanishi*, Tatsuya Nakamura*, Eli Canaani†, and Carlo M. Croce*‡ *Department of Molecular Virology, Immunology, and Medical Genetics and Comprehensive Cancer Center, Ohio State University, Columbus, OH 43210; and †Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot 76100, Israel Edited by Janet D. Rowley, University of Chicago Medical Center, Chicago, IL, and approved July 25, 2007 (received for review April 6, 2007) Erythropoietin-producing hepatoma-amplified sequence (Eph) re- studies revealed deregulation of some of the Eph/ephrin genes in ceptor tyrosine kinases and their cell-surface-bound ligands, the human malignancies, including up-regulation of ephrin-A1 or -B2 ephrins, function as a unique signaling system triggered by cell- in melanoma (2, 3), up-regulation of EphB2 in stomach cancer to-cell interaction and have been shown to mediate neurodevel- (4) and in breast cancer (5), and up-regulation of EphA2 in opmental processes. In addition, recent studies showed deregula- prostate (6), breast (7), and esophageal cancers (8), some of tion of some of Eph/ephrin genes in human malignancies, which were shown to be associated with tumor invasion or tumor suggesting the involvement of this signaling pathway in tumori- metastasis and therefore associated with poor prognosis. Con- genesis. The ALL1 (also termed MLL) gene on human chromosome versely, mutational inactivation of EphB2 was detected in pros- 11q23 was isolated by virtue of its involvement in recurrent tate (9) and colon cancers (10), suggesting tumor suppressor chromosome translocations associated with acute leukemias with function of this Eph receptor in the relevant tumors. -

The Junction-Associated Protein AF-6 Interacts and Clusters with Specific
The Junction-associated Protein AF-6 Interacts and Clusters with Specific Eph Receptor Tyrosine Kinases at Specialized Sites of Cell–Cell Contact in the Brain Michael Buchert,* Stefan Schneider,* Virginia Meskenaite,§ Mark T. Adams,* Eli Canaani,¶ Thomas Baechi,‡ Karin Moelling,* and Christopher M. Hovens* *Institut für Medizinische Virologie and ‡Elektronenmikroskopisches Zentrallabor, Universität Zürich; §Institut für Neuroinformatik, Universität Zürich/Eidgenössische Technische Hochschule, CH-8028 Zürich, Switzerland; and ¶Weizmann Institute for Science, Rehovot 76100, Israel Abstract. The AF-6/afadin protein, which contains a Eph receptors at postsynaptic membrane sites of exci- single PDZ domain, forms a peripheral component of tatory synapses in the hippocampus. Furthermore, cell membranes at specialized sites of cell–cell junc- AF-6 is a substrate for a subgroup of Eph receptors and tions. To identify potential receptor-binding targets of phosphorylation of AF-6 is dependent on a functional AF-6 we screened the PDZ domain of AF-6 against a kinase domain of the receptor. The physical interaction range of COOH-terminal peptides selected from recep- of endogenous AF-6 with Eph receptors is demon- tors having potential PDZ domain-binding termini. The strated by coimmunoprecipitation from whole rat brain PDZ domain of AF-6 interacts with a subset of mem- lysates. AF-6 is a candidate for mediating the clustering bers of the Eph subfamily of RTKs via its COOH ter- of Eph receptors at postsynaptic specializations in the minus both in vitro and in vivo. Cotransfection of a adult rat brain. green fluorescent protein-tagged AF-6 fusion protein with full-length Eph receptors into heterologous cells Key words: postsynaptic clustering • PDZ domains • induces a clustering of the Eph receptors and AF-6 at receptor tyrosine kinases • neuron physiology • Ras- sites of cell–cell contact. -

Eph Receptor Signalling: from Catalytic to Non-Catalytic Functions
Oncogene (2019) 38:6567–6584 https://doi.org/10.1038/s41388-019-0931-2 REVIEW ARTICLE Eph receptor signalling: from catalytic to non-catalytic functions 1,2 1,2 3 1,2 1,2 Lung-Yu Liang ● Onisha Patel ● Peter W. Janes ● James M. Murphy ● Isabelle S. Lucet Received: 20 March 2019 / Revised: 23 July 2019 / Accepted: 24 July 2019 / Published online: 12 August 2019 © The Author(s) 2019. This article is published with open access Abstract Eph receptors, the largest subfamily of receptor tyrosine kinases, are linked with proliferative disease, such as cancer, as a result of their deregulated expression or mutation. Unlike other tyrosine kinases that have been clinically targeted, the development of therapeutics against Eph receptors remains at a relatively early stage. The major reason is the limited understanding on the Eph receptor regulatory mechanisms at a molecular level. The complexity in understanding Eph signalling in cells arises due to following reasons: (1) Eph receptors comprise 14 members, two of which are pseudokinases, EphA10 and EphB6, with relatively uncharacterised function; (2) activation of Eph receptors results in dimerisation, oligomerisation and formation of clustered signalling centres at the plasma membrane, which can comprise different combinations of Eph receptors, leading to diverse downstream signalling outputs; (3) the non-catalytic functions of Eph receptors have been overlooked. This review provides a structural perspective of the intricate molecular mechanisms that 1234567890();,: 1234567890();,: drive Eph receptor signalling, and investigates the contribution of intra- and inter-molecular interactions between Eph receptors intracellular domains and their major binding partners. We focus on the non-catalytic functions of Eph receptors with relevance to cancer, which are further substantiated by exploring the role of the two pseudokinase Eph receptors, EphA10 and EphB6.