Fish and Tetrapods
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Early Tetrapod World; Laying the Foundations of the Modern Vertebrate Fauna
The Early Tetrapod World; laying the foundations of the modern vertebrate fauna. A one-day conference celebrating the career of Prof Jenny Clack FRS. 10.00 Welcome Head of Department of Zoology Session 1 Chair Per Ahlberg 10.10 Tim Smithson et al. Traquair’s lungfish from Loanhead: dipnoan diversity and tooth plate growth in the late Mississippian. 10.30 Mike Coates Same old fish: new name, new fins. 10.50 Zerina Johanson et al. Ontogenetic development of the otic region in the new model animal Leucoraja. 11.10 Coffee Session 2 Chair Tim Smithson 11.30 John Marshall Palynology – sometimes little things make a big difference. 11.50 Dave Millward Palaeogeography during Romer’s Gap and its potential influence on tetrapod terrestrialisation. 12.10 Sarah Davies et al. Early Carboniferous palaeoenvironments: uncovering the landscapes of Romer’s Gap. 12.30 Henning Blom New data from the late Devonian of East Greenland. 12.50 Lunch Session 3 Chair Marcello Ruta 2.00 Per Ahlberg New data, new insights and new problems: some thoughts about the origin of tetrapods. 2.20 Jason Anderson et al. Enigmatic tetrapod from Five Points, Ohio (Upper Carboniferous) further supports late survivorship of stem tetrapod lineages. 2.40 Angela Milner Keraterpeton, the earliest horned nectridean revisited. 3.10 Andrew Milner Two primitive trematopid amphibians from the Carboniferous of the Czech Republic 3.30 Tea Session 4 Chair 3.50 Nick Fraser et al. Restoring the flat-pack skull of the Triassic protorosaur Tanystropheus. 4.10 Sophie Sanchez The life history traits of stem tetrapods 4.30 Eva Herbst et al. -

A New Species of Yunnanozoan with Implications for Deuterostome
R EPORTS istry, K. S. Birdi, Ed. (CRC Press Inc., Boca Raton, FL, 23. L. H. Sperling, Polymeric Multicomponent Materials (grant EA/TU¨ BA-GEBI˙P/2001-1-1) and the Koc¸ Uni- 1997), chap. 9. (Wiley-Interscience, New York, 1997), chap. 6. versity Fiat Fund. 18. , G. McHale, M. I. Newton, Langmuir 18, 2636 24. J. Varga, in Polypropylene Structure, Blends and Com- Supporting Online Material (2002). posites, J. Karger-Kocsis, Ed. (Chapman and Hall, Lon- www.sciencemag.org/cgi/content/full/299/5611/1377/ 19. Materials and methods are available as supporting don, 1995), vol. 1, chap. 3. DC1 material on Science Online. 25. We thank A. Altay, M. A. Gu¨lgu¨n of Sabancı Univer- Materials and Methods 20. R. N. Wenzel, Ind. Eng. Chem. 28, 988 (1936). sity, and A. Alkan of Brisa for their help with SEM Figs. S1 and S2 21. A. B. D. Cassie, S. Baxter, Trans. Faraday Soc. 3, 16 (1944). measurements. A.L.D. acknowledges the financial References 22. R. A. Veselovsky, V. N. Kestelman, Adhesion of Poly- support of the Turkish Academy of Sciences in the mers (McGraw-Hill, New York, 2002), chap. 2. framework of the Young Scientist Award Program 11 September 2002; accepted 22 January 2003 and ventral units are rarely touching (Fig. 1, A New Species of Yunnanozoan G and H), suggesting that the entire anterior region could expand and contract in height. with Implications for The expansion would occur by an accommo- dation along the median zone and about an axis of rotation near the posterior of the units. -

June 2, 2016 TO
June 2, 2016 TO: Members of the Board of Regents Designated Representatives to the Board of Regents FROM: Joan Goldblatt, Secretary of the Board of Regents RE: Schedule of Meetings THURSDAY, JUNE 9, 2016 8:30 to 10:05 a.m. Petersen Room ACADEMIC AND STUDENT AFFAIRS Allen Library COMMITTEE: Regents Rice (Chair), Kritzer, Riojas, Simon *10:20 to 11:20 a.m. Petersen Room FINANCE AND ASSET MANAGEMENT Allen Library COMMITTEE: Regents Jaech (Chair), Ayer, Benoliel, Blake, Harrell 11:45 a.m. Petersen Room REGULAR MEETING OF BOARD OF Allen Library REGENTS: Regents Shanahan (Chair), Ayer, Benoliel, Blake, Harrell, Jaech, Kritzer, Rice, Riojas, Simon *or upon conclusion of the previous session. Unless otherwise indicated, committee meetings of the Board of Regents will run consecutively; starting times following the first committee are estimates only. If a session ends earlier than expected, the next scheduled session may convene immediately. Committee meetings may be attended by all members of the Board of Regents and all members may participate. To request disability accommodation, contact the Disability Services Office at: 206.543.6450 (voice), 206.543.6452 (TTY), 206.685.7264 (fax), or email at [email protected]. The University of Washington makes every effort to honor disability accommodation requests. Requests can be responded to most effectively if received as far in advance of the event as possible. 1.1/206-16 6/9/16 UNIVERSITY OF WASHINGTON BOARD OF REGENTS Academic and Student Affairs Committee Regents Rice (Chair), Kritzer, Riojas, Simon June 9, 2016 8:30 to 10:05 a.m. Petersen Room, Allen Library Approval of Minutes of Committee Meeting on May 12, 2016 COMMITTEE ACTION 1. -

O2 Secretion in the Eye and Swimbladder of Fishes
1641 The Journal of Experimental Biology 209, 1641-1652 Published by The Company of Biologists 2007 doi:10.1242/jeb.003319 Historical reconstructions of evolving physiological complexity: O2 secretion in the eye and swimbladder of fishes Michael Berenbrink School of Biological Sciences, The University of Liverpool, Biosciences Building, Crown Street, Liverpool, L69 7ZB, UK e-mail: [email protected] Accepted 12 March 2007 Summary The ability of some fishes to inflate their compressible value of haemoglobin. These changes predisposed teleost swimbladder with almost pure oxygen to maintain neutral fishes for the later evolution of swimbladder oxygen buoyancy, even against the high hydrostatic pressure secretion, which occurred at least four times independently several thousand metres below the water surface, has and can be associated with increased auditory sensitivity fascinated physiologists for more than 200·years. This and invasion of the deep sea in some groups. It is proposed review shows how evolutionary reconstruction of the that the increasing availability of molecular phylogenetic components of such a complex physiological system on a trees for evolutionary reconstructions may be as important phylogenetic tree can generate new and important insights for understanding physiological diversity in the post- into the origin of complex phenotypes that are difficult to genomic era as the increase of genomic sequence obtain with a purely mechanistic approach alone. Thus, it information in single model species. is shown that oxygen secretion first evolved in the eyes of fishes, presumably for improved oxygen supply to an Glossary available online at avascular, metabolically active retina. Evolution of this http://jeb.biologists.org/cgi/content/full/210/9/1641/DC1 system was facilitated by prior changes in the pH dependence of oxygen-binding characteristics of Key words: oxygen secretion, Root effect, rete mirabile, choroid, haemoglobin (the Root effect) and in the specific buffer swimbladder, phylogenetic reconstruction. -

Notes on the Swim-Bladder Physiology of Cod (Gadus Morhua) Investigated from the Underwater Laboratory "Helgoland"
Helgol~inder wiss. Meeresunters. 29, 460-463 (1977) Notes on the swim-bladder physiology of cod (Gadus morhua) investigated from the underwater laboratory "Helgoland" G. SUNDNES, B. GULLIKSEN ~X~ J. MORK Biological stasjon; Trondheim, Norway ABSTRACT: In situ sampling of gas from cod swim-bladders took place during a fortnight's saturation mission with the underwater laboratory "Helgoland" in May-June I975. These samples were compared to those done by the conventional method of transporting the fish to the surface for sampling. Based upon these in-situ measurements, the mean O2-concentration was 55.7 °/0 in buoyant cod at 15 m depth. Repeated sampling of the same fish showed a change in gas composition. Compared to the conventional method of transporting fish for sampling to the surface, in-situ sampling gave results with less variation, and indicated that surface-sampling does not give the correct gas composition of buoyant fish at depth of catch. INTRODUCTION Physiological investigations of fish swim-bladder have usually been performed near the surface, i.e. at about one atmosphere pressure. It means that fish were usually brought to the surface from their natural habitat of high water pressure to reduced pressure before experiments were performed and samples taken. Even in experiments with fish in pressure chambers, the gas samples of the swim-bladder were taken at one atmosphere pressure. Gas sampIes from fish at the depth of buoyancy have rarely been reported. The development of the underwater laboratory has given marine biologists a better opportunity to work under hydrostatic pressure whereby fish are kept under "natural" experimental conditions. -
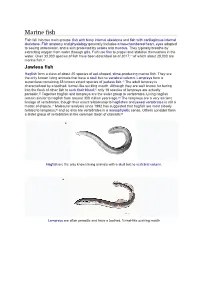
Marine Fish Fish Fall Into Two Main Groups: Fish with Bony Internal Skeletons and Fish with Cartilaginous Internal Skeletons
Marine fish Fish fall into two main groups: fish with bony internal skeletons and fish with cartilaginous internal skeletons. Fish anatomy and physiology generally includes a two-chambered heart, eyes adapted to seeing underwater, and a skin protected by scales and mucous. They typically breathe by extracting oxygen from water through gills. Fish use fins to propel and stabilise themselves in the water. Over 33,000 species of fish have been described as of 2017,[1] of which about 20,000 are marine fish.[2] Jawless fish Hagfish form a class of about 20 species of eel-shaped, slime-producing marine fish. They are the only known living animals that have a skull but no vertebral column. Lampreys form a superclass containing 38 known extant species of Jawless fish.[3] The adult lamprey is characterized by a toothed, funnel-like sucking mouth. Although they are well known for boring into the flesh of other fish to suck their blood,[4] only 18 species of lampreys are actually parasitic.[5] Together hagfish and lampreys are the sister group to vertebrates. Living hagfish remain similar to hagfish from around 300 million years ago.[6] The lampreys are a very ancient lineage of vertebrates, though their exact relationship to hagfishes and jawed vertebrates is still a matter of dispute.[7] Molecular analysis since 1992 has suggested that hagfish are most closely related to lampreys,[8] and so also are vertebrates in a monophyletic sense. Others consider them a sister group of vertebrates in the common taxon of craniata.[9] • Hagfish are the only known living animals with a skull but no vertebral column. -

Early Tetrapod Relationships Revisited
Biol. Rev. (2003), 78, pp. 251–345. f Cambridge Philosophical Society 251 DOI: 10.1017/S1464793102006103 Printed in the United Kingdom Early tetrapod relationships revisited MARCELLO RUTA1*, MICHAEL I. COATES1 and DONALD L. J. QUICKE2 1 The Department of Organismal Biology and Anatomy, The University of Chicago, 1027 East 57th Street, Chicago, IL 60637-1508, USA ([email protected]; [email protected]) 2 Department of Biology, Imperial College at Silwood Park, Ascot, Berkshire SL57PY, UK and Department of Entomology, The Natural History Museum, Cromwell Road, London SW75BD, UK ([email protected]) (Received 29 November 2001; revised 28 August 2002; accepted 2 September 2002) ABSTRACT In an attempt to investigate differences between the most widely discussed hypotheses of early tetrapod relation- ships, we assembled a new data matrix including 90 taxa coded for 319 cranial and postcranial characters. We have incorporated, where possible, original observations of numerous taxa spread throughout the major tetrapod clades. A stem-based (total-group) definition of Tetrapoda is preferred over apomorphy- and node-based (crown-group) definitions. This definition is operational, since it is based on a formal character analysis. A PAUP* search using a recently implemented version of the parsimony ratchet method yields 64 shortest trees. Differ- ences between these trees concern: (1) the internal relationships of aı¨stopods, the three selected species of which form a trichotomy; (2) the internal relationships of embolomeres, with Archeria -

Fishes Scales & Tails Scale Types 1
Phylum Chordata SUBPHYLUM VERTEBRATA Metameric chordates Linear series of cartilaginous or boney support (vertebrae) surrounding or replacing the notochord Expanded anterior portion of nervous system THE FISHES SCALES & TAILS SCALE TYPES 1. COSMOID (most primitive) First found on ostracaderm agnathans, thick & boney - composed of: Ganoine (enamel outer layer) Cosmine (thick under layer) Spongy bone Lamellar bone Perhaps selected for protection against eurypterids, but decreased flexibility 2. GANOID (primitive, still found on some living fish like gar) 3. PLACOID (old scale type found on the chondrichthyes) Dentine, tooth-like 4. CYCLOID (more recent scale type, found in modern osteichthyes) 5. CTENOID (most modern scale type, found in modern osteichthyes) TAILS HETEROCERCAL (primitive, still found on chondrichthyes) ABBREVIATED HETEROCERCAL (found on some primitive living fish like gar) DIPHYCERCAL (primitive, found on sarcopterygii) HOMOCERCAL (most modern, found on most modern osteichthyes) Agnatha (class) [connect the taxa] Cyclostomata (order) Placodermi Acanthodii (class) (class) Chondrichthyes (class) Osteichthyes (class) Actinopterygii (subclass) Sarcopterygii (subclass) Dipnoi (order) Crossopterygii (order) Ripidistia (suborder) Coelacanthiformes (suborder) Chondrostei (infra class) Holostei (infra class) Teleostei (infra class) CLASS AGNATHA ("without jaws") Most primitive - first fossils in Ordovician Bottom feeders, dorsal/ventral flattened Cosmoid scales (Ostracoderms) Pair of eyes + pineal eye - present in a few living fish and reptiles - regulates circadian rhythms Nine - seven gill pouches No paired appendages, medial nosril ORDER CYCLOSTOMATA (60 spp) Last living representatives - lampreys & hagfish Notochord not replaced by vertebrae Cartilaginous cranium, scaleless body Sea lamprey predaceous - horny teeth in buccal cavity & on tongue - secretes anti-coaggulant Lateral Line System No stomach or spleen 5 - 7 year life span - adults move into freshwater streams, spawn, & die. -

Thegreatestshowonearth-Indonesian.Pdf
1 PERTUNJUKAN TERHEBAT DI MUKA BUMI BUKTI YANG MENDUKUNG EVOLUSI RICHARD DAWKINS Diterjemahkan oleh Wahyu Ginting Terjemahan ini diterbitkan dan tersedia GRATIS di translationsproject.org 2 Daftar Isi Prakata ............................................................................................................................. 3 BAB 1 HANYA TEORI? ............................................................................................... 5 BAB 2 ANJING, SAPI, DAN KUBIS ......................................................................... 16 BAB 3 JALAN BERTABUR BUNGA MENUJU MAKRO-EVOLUSI ................. 31 BAB 4 KEHENINGAN DAN WAKTU YANG LAMBAT ...................................... 58 BAB 5 DI DEPAN MATA KEPALA KITA .............................................................. 74 BAB 6 MATA RANTAI YANG HILANG? APA MAKSUDNYA, ‘HILANG’? ... 96 BAB 7 ORANG-ORANG HILANG? TIDAK HILANG LAGI ............................. 121 BAB 8 ANDA SENDIRI MELAKUKANNYA DALAM SEMBILAN BULAN .. 139 BAB 9 BAHTERA BENUA-BENUA ....................................................................... 167 BAB 10 POHON KEKERABATAN SEPUPU ........................................................ 188 BAB 11 SEJARAH TERTULIS DI SEKUJUR TUBUH KITA ............................ 222 BAB 12 PERLOMBAAN SENJATA DAN ‘TEODISI EVOLUSIONER’ ........... 245 BAB 13 ADA KEMEGAHAN DALAM CARA PANDANG INI .......................... 260 LAMPIRAN PARA PENYANGKAL SEJARAH................................................... 279 CATATAN .................................................................................................................. -

Anatomy and Go Fish! Background
Anatomy and Go Fish! Background Introduction It is important to properly identify fi sh for many reasons: to follow the rules and regulations, for protection against sharp teeth or protruding spines, for the safety of the fi sh, and for consumption or eating purposes. When identifying fi sh, scientists and anglers use specifi c vocabulary to describe external or outside body parts. These body parts are common to most fi sh. The difference in the body parts is what helps distinguish one fi sh from another, while their similarities are used to classify them into groups. There are approximately 29,000 fi sh species in the world. In order to identify each type of fi sh, scientists have grouped them according to their outside body parts, specifi cally the number and location of fi ns, and body shape. Classifi cation Using a system of classifi cation, scientists arrange all organisms into groups based on their similarities. The fi rst system of classifi cation was proposed in 1753 by Carolus Linnaeus. Linnaeus believed that each organism should have a binomial name, genus and species, with species being the smallest organization unit of life. Using Linnaeus’ system as a guide, scientists created a hierarchical system known as taxonomic classifi cation, in which organisms are classifi ed into groups based on their similarities. This hierarchical system moves from largest and most general to smallest and most specifi c: kingdom, phylum, class, order, family, genus, and species. {See Figure 1. Taxonomic Classifi cation Pyramid}. For example, fi sh belong to the kingdom Animalia, the phylum Chordata, and from there are grouped more specifi cally into several classes, orders, families, and thousands of genus and species. -

Class Wars: Chondrichthyes and Osteichthyes Dominance in Chesapeake Bay, 2002-2012
Class Wars: Chondrichthyes and Osteichthyes dominance in Chesapeake Bay, 2002-2012. 01 July 2013 Introduction The objective of this analysis was to demonstrate a possible changing relationship between two Classes of fishes, Osteichthyes (the bony fishes) and Chondrichthyes (the cartilaginous fishes) in Chesapeake Bay based on 11 years of monitoring. If any changes between the two Classes appeared to be significant, either statistically or anecdotally, the data were explored further in an attempt to explain the variation. The Class Osteichthyes is characterized by having a skeleton made of bone and is comprised of the majority of fish species worldwide, while the Chondrichthyes skeleton is made of cartilage and is represented by the sharks, skates, and rays (the elasmobranch fishes) and chimaeras1. Many shark species are generally categorized as apex predators, while skates and rays and some smaller sharks can be placed into the mesopredator functional group (Myers et al., 2007). By definition, mesopredators prey upon a significant array of lower trophic groups, but also serve as the prey base for apex predators. Global demand for shark and consequential shark fishing mortality, estimated at 97 million sharks in 2010 (Worm et al., 2013), is hypothesized to have contributed to the decline of these apex predators in recent years (Baum et al., 2003 and Fowler et al., 2005), which in turn is suggested to have had a cascading effect on lower trophic levels—an increase in mesopredators and subsequent decrease in the prey base (Myers et al., 2007). According to 10 years of trawl survey monitoring of Chesapeake Bay, fish species composition of catches has shown a marked change over the years (Buchheister et al., 2013). -

Systematic Morphology of Fishes in the Early 21St Century
Copeia 103, No. 4, 2015, 858–873 When Tradition Meets Technology: Systematic Morphology of Fishes in the Early 21st Century Eric J. Hilton1, Nalani K. Schnell2, and Peter Konstantinidis1 Many of the primary groups of fishes currently recognized have been established through an iterative process of anatomical study and comparison of fishes that has spanned a time period approaching 500 years. In this paper we give a brief history of the systematic morphology of fishes, focusing on some of the individuals and their works from which we derive our own inspiration. We further discuss what is possible at this point in history in the anatomical study of fishes and speculate on the future of morphology used in the systematics of fishes. Beyond the collection of facts about the anatomy of fishes, morphology remains extremely relevant in the age of molecular data for at least three broad reasons: 1) new techniques for the preparation of specimens allow new data sources to be broadly compared; 2) past morphological analyses, as well as new ideas about interrelationships of fishes (based on both morphological and molecular data) provide rich sources of hypotheses to test with new morphological investigations; and 3) the use of morphological data is not limited to understanding phylogeny and evolution of fishes, but rather is of broad utility to understanding the general biology (including phenotypic adaptation, evolution, ecology, and conservation biology) of fishes. Although in some ways morphology struggles to compete with the lure of molecular data for systematic research, we see the anatomical study of fishes entering into a new and exciting phase of its history because of recent technological and methodological innovations.