Parasite-Mediated Disruptive Selection in a Natural Daphnia Population
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

And Ford, I; Ford, '953) on the Other Hand Have Put Forward a View Intermediate Between the Extreme Ones of Darwin on the One Hand and Goldschmidt on the Other
THE EVOLUTION OF MIMICRY IN THE BUTTERFLY PAPILIO DARDANUS C. A. CLARKE and P. M. SHEPPARD Departments of Medicine and Zoology, University of Liverpool Received23.V.59 1.INTRODUCTION WHENBatesputforward the mimicry hypothesis which bears his name, Darwin (1872), although accepting it, had some difficulty in explaining the evolution of the mimetic resemblance of several distinct species to one distasteful model by a series of small changes, a require- ment of his general theory of evolution. He said "it is necessary to suppose in some cases that ancient members belonging to several distinct groups, before they had diverged to their present extent, accidentally resembled a member of another and protected group in a sufficient degree to afford some slight protection; this having given the basis for the subsequent acquisition of the most perfect resemb- lance ". Punnett (1915) realised that the difficulty is even more acute when one is dealing with a polymorphic species whose forms mimic very distantly related models. Knowing that, in those butterflies which had been investigated genetically, the forms differed by single allelomorphs he concluded that the mimicry did not evolve gradually and did not confer any advantage or disadvantage to the individual. He argued that an allelomorph arises at a single step by mutation and that therefore the mimicry also arises by chance at a single step. Goldschmidt (x) although not denying that mimicry confers some advantage to its possessors also maintained that the resemblance arises fully perfected by a single mutation of a gene distinct from that producing the colour pattern in the model, but producing a similar effect in the mimic. -

Natural Selection on Phenotypes
Conner and Hartl – p. 6-1 From: Conner, J. and D. Hartl, A Primer of Ecological Genetics. In prep. for Sinauer Chapter 6: Natural selection on phenotypes Natural selection and adaptation have been recurring themes throughout this book, from the very beginning of chapter 1. We discussed selection on genotypes (and discrete phenotypes) in chapter 3, and now that we have a good understanding of the genetics of continuously distributed traits we turn to selection on these common and ecologically important phenotypes. We discuss the very general and widely used regression-based approaches to measuring selection, and cover ways to identify the phenotypic traits that are the direct targets of selection, as well as ways to determine the environmental agents that are causing selection. Identifying selective agents and targets is a powerful approach to understanding adaptation. Finally, we integrate this material with the concepts covered in chapters 4 and 5 to show how short-term phenotypic evolution can be modeled and predicted, and how this undertaking sheds light on constraints on adaptive evolution. Throughout the chapter the effects of genetic and phenotypic correlations among traits are highlighted. Evolution by natural selection has three parts (Figure 6.1; Endler 1986): 1. There is phenotypic variation for the trait of interest. 2. There is some consistent relationship between this phenotypic variation and variation in fitness. 3. A significant proportion of the phenotypic variation is caused by additive genetic variance, that is, the trait is heritable. Numbers 1 and 2 represent selection on phenotypes, which occurs within a generation and can be quantified using the selection differential (S) just as with artificial selection. -

Microevolution and the Genetics of Populations Microevolution Refers to Varieties Within a Given Type
Chapter 8: Evolution Lesson 8.3: Microevolution and the Genetics of Populations Microevolution refers to varieties within a given type. Change happens within a group, but the descendant is clearly of the same type as the ancestor. This might better be called variation, or adaptation, but the changes are "horizontal" in effect, not "vertical." Such changes might be accomplished by "natural selection," in which a trait within the present variety is selected as the best for a given set of conditions, or accomplished by "artificial selection," such as when dog breeders produce a new breed of dog. Lesson Objectives ● Distinguish what is microevolution and how it affects changes in populations. ● Define gene pool, and explain how to calculate allele frequencies. ● State the Hardy-Weinberg theorem ● Identify the five forces of evolution. Vocabulary ● adaptive radiation ● gene pool ● migration ● allele frequency ● genetic drift ● mutation ● artificial selection ● Hardy-Weinberg theorem ● natural selection ● directional selection ● macroevolution ● population genetics ● disruptive selection ● microevolution ● stabilizing selection ● gene flow Introduction Darwin knew that heritable variations are needed for evolution to occur. However, he knew nothing about Mendel’s laws of genetics. Mendel’s laws were rediscovered in the early 1900s. Only then could scientists fully understand the process of evolution. Microevolution is how individual traits within a population change over time. In order for a population to change, some things must be assumed to be true. In other words, there must be some sort of process happening that causes microevolution. The five ways alleles within a population change over time are natural selection, migration (gene flow), mating, mutations, or genetic drift. -

Patterns and Power of Phenotypic Selection in Nature
Articles Patterns and Power of Phenotypic Selection in Nature JOEL G. KINGSOLVER AND DAVID W. PFENNIG Phenotypic selection occurs when individuals with certain characteristics produce more surviving offspring than individuals with other characteristics. Although selection is regarded as the chief engine of evolutionary change, scientists have only recently begun to measure its action in the wild. These studies raise numerous questions: How strong is selection, and do different types of traits experience different patterns of selection? Is selection on traits that affect mating success as strong as selection on traits that affect survival? Does selection tend to favor larger body size, and, if so, what are its consequences? We explore these questions and discuss the pitfalls and future prospects of measuring selection in natural populations. Keywords: adaptive landscape, Cope’s rule, natural selection, rapid evolution, sexual selection henotypic selection occurs when individuals with selection on traits that affect survival stronger than on those Pdifferent characteristics (i.e., different phenotypes) that affect only mating success? In this article, we explore these differ in their survival, fecundity, or mating success. The idea and other questions about the patterns and power of phe- of phenotypic selection traces back to Darwin and Wallace notypic selection in nature. (1858), and selection is widely accepted as the primary cause of adaptive evolution within natural populations.Yet Darwin What is selection, and how does it work? never attempted to measure selection in nature, and in the Selection is the nonrandom differential survival or repro- century following the publication of On the Origin of Species duction of phenotypically different individuals. -

Arise by Chance As the Result of Mutation. They Therefore Suggest
THE EVOLUTION OF DOMINANCE UNDER DISRUPTIVE SELECTION C. A. CLARKE and P. Ni. SHEPPARD Department of Medicine and Department of Zoology, University of Liverpool Received6.iii.59 1.INTRODUCTION INa paper on the effects of disruptive selection, Mather (1955) pointed out that if there are two optimum values for a character and all others are less advantageous or disadvantageous there will be disruptive selection which can lead to the evolution of a polymorphism. Sheppard (1958) argued that where such selection is effective and the change from one optimum value to the other is switched by a single pair of allelomorphs there will be three genotypes but only two advantageous phenotypes. Consequently if dominance were absent initially it would be evolved as a result of the disruptive selection, the heterozygote and one of the homozygotes both coming to resemble one of the two optimum phenotypes (see Ford, 1955, on Tripharna comes). Thoday (1959) has shown by means of an artificial selection experiment that, even when a character is, at the beginning, controlled polygenically (sternopleural chaeta-number in Drosophila) and there is 50 per cent. gene exchange between the "high" and "low" selected sub-popu- lations, a polymorphism can evolve. The most fully understood examples of disruptive selection (other than sex) are provided by instances of Batesian Mimicry, where there are a number of distinct warningly coloured species, acting as models, which are mimicked by the polymorphic forms of a single more edible species. Fisher and Ford (see Ford, 1953) have argued that a suffi- ciently good resemblance between mimic and model is not likely to arise by chance as the result of mutation. -
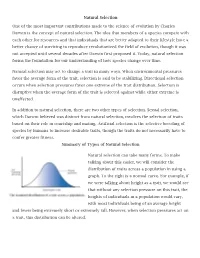
Natural Selection One of the Most Important Contributions Made to The
Natural Selection One of the most important contributions made to the science of evolution by Charles Darwin is the concept of natural selection. The idea that members of a species compete with each other for resources and that individuals that are better adapted to their lifestyle have a better chance of surviving to reproduce revolutionized the field of evolution, though it was not accepted until several decades after Darwin first proposed it. Today, natural selection forms the foundation for our understanding of how species change over time. Natural selection may act to change a trait in many ways. When environmental pressures favor the average form of the trait, selection is said to be stabilizing. Directional selection occurs when selection pressures favor one extreme of the trait distribution. Selection is disruptive when the average form of the trait is selected against while either extreme is unaffected. In addition to natural selection, there are two other types of selection. Sexual selection, which Darwin believed was distinct from natural selection, involves the selection of traits based on their role in courtship and mating. Artificial selection is the selective breeding of species by humans to increase desirable traits, though the traits do not necessarily have to confer greater fitness. Summary of Types of Natural Selection Natural selection can take many forms. To make talking about this easier, we will consider the distribution of traits across a population in using a graph. To the right is a normal curve. For example, if we were talking about height as a trait, we would see that without any selection pressure on this trait, the heights of individuals in a population would vary, with most individuals being of an average height and fewer being extremely short or extremely tall. -

Mathematics and Biology Have Not Been Easy Bedfellows and There Are
Mathematics and biology have not been easy bedfellows and there are many ex- amples of biologists who have made terrible mistakes by ignoring mathematics and also mathematicians who have not fared very well when they have dipped their toes in the biological pool. Russ’s work is a great example of the fruitful interaction between the two fi elds and it gives me enormous pleasure to call on him to deliver his lecture. Russell Lande Theoretical Population Biology: (A) Evolution of correlated characters (B) Stochastic demography and conservation (A) Evolution of correlated characters: The fundamental principle of natural selection by which living organisms adapt to their environments was discovered by Darwin (1859), who integrated a wide variety of evidence supporting the theory of evolution. Differences in individual fi tness due to variation among individuals in characters infl uencing survival and reproduction, combined with partial heritability of the variation, causes adaptive evolution. The validity of this theory is strongly confi rmed by modern genetic data. Darwin and the early naturalists understood that natural selection acts simultaneously on many characters of an organism, and that hereditary constraints among characters prevent their independent evolution. On the Origin of Species, Darwin (1859) wrote that when man or nature selects on a given character, that character as well as other correlated characters evolve, due to ‘correla- tion of growth’. We now know that genetic correlations between characters are caused by pleiotropy (a single gene infl uencing multiple characters) and linkage disequilib- rium (nonrandom association between allelic forms of different genes, especially on the same chromosome). -

Consequences of the Domestication of Man's Best Friend, The
Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 289 Consequences of the Domestication of Man’s Best Friend, The Dog SUSANNE BJÖRNERFELDT ACTA UNIVERSITATIS UPSALIENSIS ISSN 1651-6214 UPPSALA ISBN 978-91-554-6854-5 2007 urn:nbn:se:uu:diva-7799 ! "# $ % &''( '!)'' * # * * +# #, "# - #, ./ * 0, &''(, 1 * # * $ 2 3 "# , 4 , &5!, 67 , , 809 !(5:!;:<<7:65<7:<, "# - # * # ;< ''' , " # # - # # = # , # - * # - * * # * * #, "# # * # # # # # 2 # # # # , " # - # - ) # # 94 # > # # , 8 1 # - # ** - # # # 2 * , 3 - * # - # # -# * , "# # * # # # * * * # # # , ? # * , * 1 * * - # *- * , @ - * # * - # -## * * , "# 1 # # # * * # * * , + 2 * # * - # - - # 2 , # > # -# * * - 1 * !" # $ # % # $ # & '( )*# # $+,-./0 # A 0 ./ * &''( 8009 ;6<;:6&;7 809 !(5:!;:<<7:65<7:< ) ))) :((!! B# )CC ,,C D E ) ))) :((!!F All knowledge, the totality of all questions and answers, is contained in the dog. FRANZ KAFKA, Investigations of the dog Cover photos by S Björnerfeldt List of Papers This thesis -

Biol2007 - Evolution of Quantitative Characters
1 BIOL2007 - EVOLUTION OF QUANTITATIVE CHARACTERS How do evolutionary biologists measure variation in a typical quantitative character? Let’s use beak size in birds as a typical example. Phenotypic variation (Vp) in a quantitative character Some of the variation in beak size is caused by environmental factors (VE). Some of the variation is caused by genetic factors (VG). If there was no variation in genotype between birds then individuals would differ in beak size only because of different rearing environments. Quantitative geneticists think in terms of how the value of a character in an individual compares to the mean value of the character in the population. The population mean is taken to be a neutral measure and then particular phenotypes are increases or decreases from that value. AT LEVEL OF INDIVIDUAL Phenotype = genotype + environment P = G + E AT LEVEL OF POPULATION Phenotypic variance = genotypic variance + environmental variance VP = VG + VE Genotypic variance is divided into additive, dominance and interaction components VG = VA + VD + V I VA = Additive variance – due to the additive effects of individual alleles (inherited) VD = Dominance variance - reflect the combinations of alleles at each locus (homo- versus (homozygotes versus hetero-zygotes) in any given generation but are not inherited by offspring. An intra-locus effect. VI = Interaction/epistatic variance - reflect epistatic interactions between alleles at different loci. Again not passed onto offspring; they depend on particular combinations of genes in a given generation and are not inherited by offspring. An inter-locus effect. ___________________________________________________________ The following section (italicised) provides the alebgra to backup my assertion that VD and VI are not heritable. -

FULLTEXT01.Pdf
Journal of Theoretical Biology 507 (2020) 110449 Contents lists available at ScienceDirect Journal of Theoretical Biology journal homepage: www.elsevier.com/locate/yjtbi The components of directional and disruptive selection in heterogeneous group-structured populations ⇑ Hisashi Ohtsuki a, , Claus Rueffler b, Joe Yuichiro Wakano c,d,e, Kalle Parvinen f,g, Laurent Lehmann h a Department of Evolutionary Studies of Biosystems, School of Advanced Sciences, SOKENDAI (The Graduate University for Advanced Studies), Shonan Village, Hayama, Kanagawa 240-0193, Japan b Animal Ecology, Department of Ecology and Genetics, University of Uppsala, Sweden c School of Interdisciplinary Mathematical Sciences, Meiji University, Tokyo 164-8525, Japan d Meiji Institute for Advanced Study of Mathematical Sciences, Tokyo 164-8525, Japan e Department of Mathematics, The University of British Columbia, 1984 Mathematics Road, Vancouver, B.C. V6T 1Z2, Canada f Department of Mathematics and Statistics, FI-20014 University of Turku, Finland g Evolution and Ecology Program, International Institute for Applied Systems Analysis (IIASA), A-2361 Laxenburg, Austria h Department of Ecology and Evolution, University of Lausanne, Switzerland article info abstract Article history: We derive how directional and disruptive selection operate on scalar traits in a heterogeneous group- Received 31 March 2020 structured population for a general class of models. In particular, we assume that each group in the Revised 5 August 2020 population can be in one of a finite number of states, where states can affect group size and/or other envi- Accepted 7 August 2020 ronmental variables, at a given time. Using up to second-order perturbation expansions of the invasion Available online 16 August 2020 fitness of a mutant allele, we derive expressions for the directional and disruptive selection coefficients, which are sufficient to classify the singular strategies of adaptive dynamics. -

Predator Mediated Selection and the Impact of Developmental Stage on Viability in Wood Frog Tadpoles (Rana Sylvatica) Ryan Calsbeek1,2* and Shawn Kuchta1,3
Calsbeek and Kuchta BMC Evolutionary Biology 2011, 11:353 http://www.biomedcentral.com/1471-2148/11/353 RESEARCHARTICLE Open Access Predator mediated selection and the impact of developmental stage on viability in wood frog tadpoles (Rana sylvatica) Ryan Calsbeek1,2* and Shawn Kuchta1,3 Abstract Background: Complex life histories require adaptation of a single organism for multiple ecological niches. Transitions between life stages, however, may expose individuals to an increased risk of mortality, as the process of metamorphosis typically includes developmental stages that function relatively poorly in both the pre- and post- metamorphic habitat. We studied predator-mediated selection on tadpoles of the wood frog, Rana sylvatica,to identify this hypothesized period of differential predation risk and estimate its ontogenetic onset. We reared tadpoles in replicated mesocosms in the presence of the larval odonate Anax junius, a known tadpole predator. Results: The probability of tadpole survival increased with increasing age and size, but declined steeply at the point in development where hind limbs began to erupt from the body wall. Selection gradient analyses indicate that natural selection favored tadpoles with short, deep tail fins. Tadpoles resorb their tails as they progress toward metamorphosis, which may have led to the observed decrease in survivorship. Path models revealed that selection acted directly on tail morphology, rather than through its indirect influence on swimming performance. Conclusions: This is consistent with the hypothesis that tail morphology influences predation rates by reducing the probability a predator strikes the head or body. Background among different life stages allow individuals to exploit Many organisms exploit different environments over the multiple kinds of resources throughout ontogeny, com- course of their life cycle. -

Mate Choice and Sexual Selection: What Have We Learned Since Darwin?
Mate choice and sexual selection: What have we learned since Darwin? Adam G. Jones1 and Nicholas L. Ratterman Department of Biology, Texas A&M University, 3258 TAMU, College Station, TX 77843 Charles Darwin laid the foundation for all modern work on sexual concerns sexual selection, but many of Darwin’s insights regard- selection in his seminal book The Descent of Man, and Selection in ing sexual selection appear in his chapters on humans. Relation to Sex. In this work, Darwin fleshed out the mechanism of Darwin’s most lasting achievement with respect to sexual sexual selection, a hypothesis that he had proposed in The Origin of selection must be his definition of the term, as it is essentially the Species. He went well beyond a simple description of the phenom- same as the one still in use today. It is difficult to find a quote enon by providing extensive evidence and considering the far-reach- from Darwin that captures the full essence of his concept of ing implications of the idea. Here we consider the contributions of sexual selection, but he provides the following definition (ref. 2; Darwin to sexual selection with a particular eye on how far we have Part I, pp 254–255): progressed in the last 150 years. We focus on 2 key questions in sexual selection. First, why does mate choice evolve at all? And second, what ‘‘We are, however, here concerned only with that kind factors determine the strength of mate choice (or intensity of sexual of selection, which I have called sexual selection. This selection) in each sex? Darwin provided partial answers to these depends on the advantage which certain individuals questions, and the progress that has been made on both of these have over other individuals of the same sex and species, topics since his time should be seen as one of the great triumphs of in exclusive relation to reproduction.’’ modern evolutionary biology.