On the Growth of Polytrichum, Pleurozium
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Coptis Trifolia Conservation Assessment
CONSERVATION ASSESSMENT for Coptis trifolia (L.) Salisb. Originally issued as Management Recommendations December 1998 Marty Stein Reconfigured-January 2005 Tracy L. Fuentes USDA Forest Service Region 6 and USDI Bureau of Land Management, Oregon and Washington CONSERVATION ASSESSMENT FOR COPTIS TRIFOLIA Table of Contents Page List of Tables ................................................................................................................................. 2 List of Figures ................................................................................................................................ 2 Summary........................................................................................................................................ 4 I. NATURAL HISTORY............................................................................................................. 6 A. Taxonomy and Nomenclature.......................................................................................... 6 B. Species Description ........................................................................................................... 6 1. Morphology ................................................................................................................... 6 2. Reproductive Biology.................................................................................................... 7 3. Ecological Roles ............................................................................................................. 7 C. Range and Sites -

Economic and Ethnic Uses of Bryophytes
Economic and Ethnic Uses of Bryophytes Janice M. Glime Introduction Several attempts have been made to persuade geologists to use bryophytes for mineral prospecting. A general lack of commercial value, small size, and R. R. Brooks (1972) recommended bryophytes as guides inconspicuous place in the ecosystem have made the to mineralization, and D. C. Smith (1976) subsequently bryophytes appear to be of no use to most people. found good correlation between metal distribution in However, Stone Age people living in what is now mosses and that of stream sediments. Smith felt that Germany once collected the moss Neckera crispa bryophytes could solve three difficulties that are often (G. Grosse-Brauckmann 1979). Other scattered bits of associated with stream sediment sampling: shortage of evidence suggest a variety of uses by various cultures sediments, shortage of water for wet sieving, and shortage around the world (J. M. Glime and D. Saxena 1991). of time for adequate sampling of areas with difficult Now, contemporary plant scientists are considering access. By using bryophytes as mineral concentrators, bryophytes as sources of genes for modifying crop plants samples from numerous small streams in an area could to withstand the physiological stresses of the modern be pooled to provide sufficient material for analysis. world. This is ironic since numerous secondary compounds Subsequently, H. T. Shacklette (1984) suggested using make bryophytes unpalatable to most discriminating tastes, bryophytes for aquatic prospecting. With the exception and their nutritional value is questionable. of copper mosses (K. G. Limpricht [1885–]1890–1903, vol. 3), there is little evidence of there being good species to serve as indicators for specific minerals. -

Vermont Natural Community Types
Synonymy of Vermont Natural Community Types with National Vegetation Classification Associations Eric Sorenson and Bob Zaino Natural Heritage Inventory Vermont Fish and Wildlife Department October 17, 2019 Vermont Natural Community Type Patch State National and International Vegetation Classification. NatureServe. 2019. Size Rank NatureServe Explorer: An online encyclopedia of life. NatureServe, Arlington, Virginia. Spruce-Fir-Northern Hardwood Forest Formation Subalpine Krummholz S S1 Picea mariana - Abies balsamea / Sibbaldiopsis tridentata Shrubland (CEGL006038); (Picea mariana, Abies balsamea) / Kalmia angustifolia - Ledum groenlandicum Dwarf-shrubland Montane Spruce-Fir Forest L-M S3 Picea rubens - Abies balsamea - Sorbus americana Forest (CEGL006128) Variant: Montane Fir Forest L-M S3 Abies balsamea - (Betula papyrifera var. cordifolia) Forest (CEGL006112) Variant: Montane Spruce Forest Lowland Spruce-Fir Forest L-M S3 Picea mariana - Picea rubens / Pleurozium schreberi Forest (CEGL006361) Variant: Well-Drained Lowland Spruce- L S2 Picea rubens - Abies balsamea - Betula papyrifera Forest (CEGL006273); Fir Forest Picea mariana - Picea rubens / Rhododendron canadense / Cladina spp. Woodland (CEGL006421) Montane Yellow Birch-Red Spruce Forest M S3 Betula alleghaniensis - Picea rubens / Dryopteris campyloptera Forest (CEGL006267) Variant: Montane Yellow Birch-Sugar L S3 Maple-Red Spruce Forest Red Spruce-Northern Hardwood Forest M S5 Betula alleghaniensis - Picea rubens / Dryopteris campyloptera Forest (CEGL006267) Red Spruce-Heath -

Appendix 2: Plant Lists
Appendix 2: Plant Lists Master List and Section Lists Mahlon Dickerson Reservation Botanical Survey and Stewardship Assessment Wild Ridge Plants, LLC 2015 2015 MASTER PLANT LIST MAHLON DICKERSON RESERVATION SCIENTIFIC NAME NATIVENESS S-RANK CC PLANT HABIT # OF SECTIONS Acalypha rhomboidea Native 1 Forb 9 Acer palmatum Invasive 0 Tree 1 Acer pensylvanicum Native 7 Tree 2 Acer platanoides Invasive 0 Tree 4 Acer rubrum Native 3 Tree 27 Acer saccharum Native 5 Tree 24 Achillea millefolium Native 0 Forb 18 Acorus calamus Alien 0 Forb 1 Actaea pachypoda Native 5 Forb 10 Adiantum pedatum Native 7 Fern 7 Ageratina altissima v. altissima Native 3 Forb 23 Agrimonia gryposepala Native 4 Forb 4 Agrostis canina Alien 0 Graminoid 2 Agrostis gigantea Alien 0 Graminoid 8 Agrostis hyemalis Native 2 Graminoid 3 Agrostis perennans Native 5 Graminoid 18 Agrostis stolonifera Invasive 0 Graminoid 3 Ailanthus altissima Invasive 0 Tree 8 Ajuga reptans Invasive 0 Forb 3 Alisma subcordatum Native 3 Forb 3 Alliaria petiolata Invasive 0 Forb 17 Allium tricoccum Native 8 Forb 3 Allium vineale Alien 0 Forb 2 Alnus incana ssp rugosa Native 6 Shrub 5 Alnus serrulata Native 4 Shrub 3 Ambrosia artemisiifolia Native 0 Forb 14 Amelanchier arborea Native 7 Tree 26 Amphicarpaea bracteata Native 4 Vine, herbaceous 18 2015 MASTER PLANT LIST MAHLON DICKERSON RESERVATION SCIENTIFIC NAME NATIVENESS S-RANK CC PLANT HABIT # OF SECTIONS Anagallis arvensis Alien 0 Forb 4 Anaphalis margaritacea Native 2 Forb 3 Andropogon gerardii Native 4 Graminoid 1 Andropogon virginicus Native 2 Graminoid 1 Anemone americana Native 9 Forb 6 Anemone quinquefolia Native 7 Forb 13 Anemone virginiana Native 4 Forb 5 Antennaria neglecta Native 2 Forb 2 Antennaria neodioica ssp. -

Molecular Phylogeny of Chinese Thuidiaceae with Emphasis on Thuidium and Pelekium
Molecular Phylogeny of Chinese Thuidiaceae with emphasis on Thuidium and Pelekium QI-YING, CAI1, 2, BI-CAI, GUAN2, GANG, GE2, YAN-MING, FANG 1 1 College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China. 2 College of Life Science, Nanchang University, 330031 Nanchang, China. E-mail: [email protected] Abstract We present molecular phylogenetic investigation of Thuidiaceae, especially on Thudium and Pelekium. Three chloroplast sequences (trnL-F, rps4, and atpB-rbcL) and one nuclear sequence (ITS) were analyzed. Data partitions were analyzed separately and in combination by employing MP (maximum parsimony) and Bayesian methods. The influence of data conflict in combined analyses was further explored by two methods: the incongruence length difference (ILD) test and the partition addition bootstrap alteration approach (PABA). Based on the results, ITS 1& 2 had crucial effect in phylogenetic reconstruction in this study, and more chloroplast sequences should be combinated into the analyses since their stability for reconstructing within genus of pleurocarpous mosses. We supported that Helodiaceae including Actinothuidium, Bryochenea, and Helodium still attributed to Thuidiaceae, and the monophyletic Thuidiaceae s. lat. should also include several genera (or species) from Leskeaceae such as Haplocladium and Leskea. In the Thuidiaceae, Thuidium and Pelekium were resolved as two monophyletic groups separately. The results from molecular phylogeny were supported by the crucial morphological characters in Thuidiaceae s. lat., Thuidium and Pelekium. Key words: Thuidiaceae, Thuidium, Pelekium, molecular phylogeny, cpDNA, ITS, PABA approach Introduction Pleurocarpous mosses consist of around 5000 species that are defined by the presence of lateral perichaetia along the gametophyte stems. Monophyletic pleurocarpous mosses were resolved as three orders: Ptychomniales, Hypnales, and Hookeriales (Shaw et al. -

Genetic Variation in the Clonal Bryophyte Hylocomium Splendens at Hierarchical Geographical Scales in Scandinavia
Heredity 78 (1997) 293-301 Received 26 April 1996 Genetic variation in the clonal bryophyte Hylocomium splendens at hierarchical geographical scales in Scandinavia NILS CRONBERG*t, ULF MOLAU:j: & MATS SONESSON§ tDepartment of Systematic Botany, University of Lund, 6. Val/gatan 18-20, S-223 61 Lund, tDepartment of Systematic Botany, University of Goteborg, Carl Skottsbergs Gata 22, S-413 19 Goteborg and §Abisko Scientific Research Station, The Royal Swedish Academy of Sciences, S-981 07 Abisko, Sweden The widespread bryophyte Hylocomium splendens was sampled in a hierarchical fashion from populations representing four Scandinavian vegetation zones. Allozyme electrophoresis revealed variation at 11 out of 13 screened loci, allowing accurate identification of genotypes. From a total sample of 298 shoots 79 genotypes could be detected, giving the proportion of distinguishable genotypes (PD) of 0.265. The total allelic diversity (HT) based on polymorphic loci was 0.274. The relative differentiation among populations was low (GsT = 0.073), indicating a high level of gene flow. Differences in population structuring occurred between a subarctic alpine site vs. three lowland sites. The subarctic-alpine population had one widespread clone, which appeared to be propagated by dispersal of vegetative fragments. That population also comprised many rare genotypes, often occurring together within 10 x 10 cm patches. The lowland populations had genotypes that were locally common and often dominant within the patches. When identical genotypes were observed in multiple patches within these populations, it was usually statistically highly probable that they had arisen by independent sexual recombinations. Keywords: allozyme, Bryophyta, clonal plant, genetic diversity, hierarchical variation, Hy/oco mium splendens. -
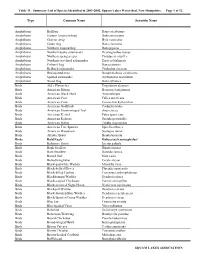
Summary Data
Table 11. Summary List of Species Identified in 2001-2002, Squam Lakes Watershed, New Hampshire. Page 1 of 12 Type Common Name Scientific Name Amphibians Bullfrog Rana catesbeiana Amphibians Eastern American toad Bufo americanus Amphibians Gray treefrog Hyla versicolor Amphibians Green frog Rana clamitans Amphibians Northern leopard frog Rana pipiens Amphibians Northern dusky salamander Desmognathus fuscus Amphibians Northern spring peeper Pseudacris crucifer Amphibians Northern two-lined salamander Eurycea bislineata Amphibians Pickerel frog Rana palustris Amphibians Redback salamander Plethodon cinereus Amphibians Red-spotted newt Notophthalmus viridescens Amphibians Spotted salamander Ambystoma maculatum Amphibians Wood frog Rana sylvatica Birds Alder Flycatcher Empidonax alnorum Birds American Bittern Botaurus lentiginosus Birds American Black Duck Anas rubripes Birds American Coot Fulica americana Birds American Crow Corvus brachyrhynchos Birds American Goldfinch Carduelis tristis Birds American Green-winged Teal Anas crecca Birds American Kestrel Falco sparverius Birds American Redstart Setophaga ruticilla Birds American Robin Turdus migratorius Birds American Tree Sparrow Spizella arborea Birds American Woodcock Scolopax minor Birds Atlantic Brant Branta bernicla Birds Bald Eagle* Haliaeetus leucocephalus* Birds Baltimore Oriole Icterus galbula Birds Bank Swallow Riparia riparia Birds Barn Swallow Hirundo rustica Birds Barred Owl Strix varia Birds Belted Kingfisher Ceryle alcyon Birds Black-and-white Warbler Mniotilta varia Birds -

Volume 1, Chapter 2-7: Bryophyta
Glime, J. M. 2017. Bryophyta – Bryopsida. Chapt. 2-7. In: Glime, J. M. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook 2-7-1 sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 10 January 2019 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology/>. CHAPTER 2-7 BRYOPHYTA – BRYOPSIDA TABLE OF CONTENTS Bryopsida Definition........................................................................................................................................... 2-7-2 Chromosome Numbers........................................................................................................................................ 2-7-3 Spore Production and Protonemata ..................................................................................................................... 2-7-3 Gametophyte Buds.............................................................................................................................................. 2-7-4 Gametophores ..................................................................................................................................................... 2-7-4 Location of Sex Organs....................................................................................................................................... 2-7-6 Sperm Dispersal .................................................................................................................................................. 2-7-7 Release of Sperm from the Antheridium..................................................................................................... -

Here Is Glaucum) Looks Like Tiny Pine Seedlings on This Disturbed Soil Along Forest Edge
Pennsylvania Natural Heritage Program informationinformation forfor thethe conservationconservation ofof biodiversitybiodiversity Wild Heritage News January—March 2014 Bryophytes and Lichens of Dry Oak Heath Forest Communities Inside This Issue by Scott Schuette Bryophytes and Pg 1 Lichens Pennsylvania is flush with a diversity of biodiversity ripe for exploration if one just Edge of Range Pg 4 forest communities, some garnering looks a little bit closer at the smaller Species special attention due to the quality of organisms living within this forest plant diversity and others for the community. Preserving Records Pg 6 hardwood resources they provide. The of our Work dry oak heath forests fall into the latter Bryophytes and lichens are present in category. These forest communities are every forest community, regardless of size Notes from the Pg 7 found throughout the state occurring on and condition. It is well-documented that Field moderately dry to xeric, acidic sites that bryophytes and lichens are sensitive to Measures of Pg 12 usually have shallow or sometimes sandy human disturbance and, as a result, have Progress soils. The dominant tree canopy is been used as indicators of habitat quality. chestnut oak (Quercus montana) mixed These organisms tend to live on the trees with black oak (Quercus velutina), white and shrubs as epiphytes (rootless, oak (Quercus alba), and red maple (Acer independent living plants), but they can rubrum). Other tree species that are also completely cover boulders or reside commonly present in the subcanopy Photo Banner: include sassafras (Sassafras albidum), Scott Schuette sweet birch (Betula lenta), black gum (Nyssa sylvatica), and eastern white pine The stair-step moss (Pinus strobus). -

Kenai National Wildlife Refuge Species List, Version 2018-07-24
Kenai National Wildlife Refuge Species List, version 2018-07-24 Kenai National Wildlife Refuge biology staff July 24, 2018 2 Cover image: map of 16,213 georeferenced occurrence records included in the checklist. Contents Contents 3 Introduction 5 Purpose............................................................ 5 About the list......................................................... 5 Acknowledgments....................................................... 5 Native species 7 Vertebrates .......................................................... 7 Invertebrates ......................................................... 55 Vascular Plants........................................................ 91 Bryophytes ..........................................................164 Other Plants .........................................................171 Chromista...........................................................171 Fungi .............................................................173 Protozoans ..........................................................186 Non-native species 187 Vertebrates ..........................................................187 Invertebrates .........................................................187 Vascular Plants........................................................190 Extirpated species 207 Vertebrates ..........................................................207 Vascular Plants........................................................207 Change log 211 References 213 Index 215 3 Introduction Purpose to avoid implying -

Species List For: Labarque Creek CA 750 Species Jefferson County Date Participants Location 4/19/2006 Nels Holmberg Plant Survey
Species List for: LaBarque Creek CA 750 Species Jefferson County Date Participants Location 4/19/2006 Nels Holmberg Plant Survey 5/15/2006 Nels Holmberg Plant Survey 5/16/2006 Nels Holmberg, George Yatskievych, and Rex Plant Survey Hill 5/22/2006 Nels Holmberg and WGNSS Botany Group Plant Survey 5/6/2006 Nels Holmberg Plant Survey Multiple Visits Nels Holmberg, John Atwood and Others LaBarque Creek Watershed - Bryophytes Bryophte List compiled by Nels Holmberg Multiple Visits Nels Holmberg and Many WGNSS and MONPS LaBarque Creek Watershed - Vascular Plants visits from 2005 to 2016 Vascular Plant List compiled by Nels Holmberg Species Name (Synonym) Common Name Family COFC COFW Acalypha monococca (A. gracilescens var. monococca) one-seeded mercury Euphorbiaceae 3 5 Acalypha rhomboidea rhombic copperleaf Euphorbiaceae 1 3 Acalypha virginica Virginia copperleaf Euphorbiaceae 2 3 Acer negundo var. undetermined box elder Sapindaceae 1 0 Acer rubrum var. undetermined red maple Sapindaceae 5 0 Acer saccharinum silver maple Sapindaceae 2 -3 Acer saccharum var. undetermined sugar maple Sapindaceae 5 3 Achillea millefolium yarrow Asteraceae/Anthemideae 1 3 Actaea pachypoda white baneberry Ranunculaceae 8 5 Adiantum pedatum var. pedatum northern maidenhair fern Pteridaceae Fern/Ally 6 1 Agalinis gattingeri (Gerardia) rough-stemmed gerardia Orobanchaceae 7 5 Agalinis tenuifolia (Gerardia, A. tenuifolia var. common gerardia Orobanchaceae 4 -3 macrophylla) Ageratina altissima var. altissima (Eupatorium rugosum) white snakeroot Asteraceae/Eupatorieae 2 3 Agrimonia parviflora swamp agrimony Rosaceae 5 -1 Agrimonia pubescens downy agrimony Rosaceae 4 5 Agrimonia rostellata woodland agrimony Rosaceae 4 3 Agrostis elliottiana awned bent grass Poaceae/Aveneae 3 5 * Agrostis gigantea redtop Poaceae/Aveneae 0 -3 Agrostis perennans upland bent Poaceae/Aveneae 3 1 Allium canadense var. -

Atlantic Woodlands in the Lake District Mosses & Liverworts
LIVERWORTS generally have two rows of leaves, one up each side of the stem We are Plantlife Further information For over 25 years, Plantlife has had a single ideal Bazzania trilobata Greater whipwort Plagiochila spinulosa Prickly featherwort – to save and celebrate wild flowers, plants and Books fungi. They are the life support for all our wildlife Mosses and Liverworts of Britain and Ireland: A Field Guide 3 and their colour and character light up our by Ian Atherton, Sam Bosanquet and Mark Lawley (British landscapes. But without our help, this priceless Bryological Society, 2010). The best field guide. 2 natural heritage is in danger of being lost. The Liverwort Flora of the British Isles by Jean Paton (Harley 3 Books, 1999). An in-depth guide to liverwort identification, From the open spaces of our nature reserves to for more advanced bryologists. the corridors of government, we work nationally The Moss Flora of Britain and Ireland by A J E Smith and internationally to raise their profile, (2nd edition, Cambridge University Press, 2004). 1 celebrate their beauty and to protect their future. An in-depth guide to liverwort identification, for more advanced bryologists. 2 Mosses and Liverworts by Ron Porley and Nick Hodgetts Where wild plants lead (Collins New Naturalist series, Harper Collins, 2005). Habitat Often forming dense, mounded colonies on earth banks and around tree bases; also on boulders, rotten wood and Habitat In loose mats or dense cushions on trees and rock faces, often close to rivers and streams. Wildlife follows A highly readable account of moss and liverwort ecology occasionally on tree trunks.