Breeding Distribution of Dunlin <I>Calidris Alpina</I> in Russia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Regional Maps of Locations Mentioned in Global Review of The
Regional Maps of Locations Mentioned in Global Review of the Conservation Status of Monodontid Stocks These maps provide the locations of the geographic features mentioned in the Global Review of the Conservation Status of Monodontid Stocks. Figure 1. Locations associated with beluga stocks of the Okhotsk Sea (beluga stocks 1-5). Numbered locations are: (1) Amur River, (2) Ul- bansky Bay, (3) Tugursky Bay, (4) Udskaya Bay, (5) Nikolaya Bay, (6) Ulban River, (7) Big Shantar Island, (8) Uda River, (9) Torom River. Figure 2. Locations associated with beluga stocks of the Bering Sea and Gulf of Alaska (beluga stocks 6-9). Numbered locations are: (1) Anadyr River Estuary, (2) Anadyr River, (3) Anadyr City, (4) Kresta Bay, (5) Cape Navarin, (6) Yakutat Bay, (7) Knik Arm, (8) Turnagain Arm, (9) Anchorage, (10) Nushagak Bay, (11) Kvichak Bay, (12) Yukon River, (13) Kuskokwim River, (14) Saint Matthew Island, (15) Round Island, (16) St. Lawrence Island. Figure 3. Locations associated with beluga stocks of the Chukchi and Beaufort Seas, Canadian Arctic and West Greenland (beluga stocks 10-12 and 19). Numbered locations are: (1) St. Lawrence Island, (2) Kotzebue Sound, (3) Kasegaluk Lagoon, (4) Point Lay, (5) Wain- wright, (6) Mackenzie River, (7) Somerset Island, (8) Radstock Bay, (9) Maxwell Bay, (10) Croker Bay, (11) Devon Island, (12) Cunning- ham Inlet, (13) Creswell Bay, (14) Mary River Mine, (15) Elwin Bay, (16) Coningham Bay, (17) Prince of Wales Island, (18) Qeqertarsuat- siaat, (19) Nuuk, (20) Maniitsoq, (21) Godthåb Fjord, (22) Uummannaq, (23) Upernavik. Figure 4. Locations associated with beluga stocks of subarctic eastern Canada, Hudson Bay, Ungava Bay, Cumberland Sound and St. -

Sources and Pathways 4.1
Chapter 4 Persistant toxic substances (PTS) sources and pathways 4.1. Introduction Chapter 4 4.1. Introduction 4.2. Assessment of distant sources: In general, the human environment is a combination Longrange atmospheric transport of the physical, chemical, biological, social and cultur- Due to the nature of atmospheric circulation, emission al factors that affect human health. It should be recog- sources located within the Northern Hemisphere, par- nized that exposure of humans to PTS can, to certain ticularly those in Europe and Asia, play a dominant extent, be dependant on each of these factors. The pre- role in the contamination of the Arctic. Given the spa- cise role differs depending on the contaminant con- tial distribution of PTS emission sources, and their cerned, however, with respect to human intake, the potential for ‘global’ transport, evaluation of long- chain consisting of ‘source – pathway – biological avail- range atmospheric transport of PTS to the Arctic ability’ applies to all contaminants. Leaving aside the region necessarily involves modeling on the hemi- biological aspect of the problem, this chapter focuses spheric/global scale using a multi-compartment on PTS sources, and their physical transport pathways. approach. To meet these requirements, appropriate modeling tools have been developed. Contaminant sources can be provisionally separated into three categories: Extensive efforts were made in the collection and • Distant sources: Located far from receptor sites in preparation of input data for modeling. This included the Arctic. Contaminants can reach receptor areas the required meteorological and geophysical informa- via air currents, riverine flow, and ocean currents. tion, and data on the physical and chemical properties During their transport, contaminants are affected by of both the selected substances and of their emissions. -

CRYOSTRATIGRAPHY of the FIRST TERRACE in BELY ISLAND, KARA SEA: PERMAFROST and CLIMATE HISTORY (Part 3) E.A
Kriosfera Zemli, 2014, vol. XVIII, No. 3, pp. 32–43 http://www.izdatgeo.ru CRYOSTRATIGRAPHY OF THE FIRST TERRACE IN BELY ISLAND, KARA SEA: PERMAFROST AND CLIMATE HISTORY (Part 3) E.A. Slagoda1,2, A.N. Kurchatova1,2, O.L. Opokina1,2, I.V. Tomberg3, T.V. Khodzher3, A.D. Firsova3, E.V. Rodionova3, K.A. Popov1, E.L. Nikulina1 1 Earth Cryosphere Institute, Siberian Branch of the Russian Academy of Sciences, P/O box 1230 Tyumen, 625000, Russia; [email protected] 2Tyumen State Oil and Gas University, 56 Volodarskogo str., Tyumen, 625000, Russia 3Limnological Institute, Siberian Branch of the Russian Academy of Sciences, 3 Ulan-Batorskaya str., Irkutsk, 664033, Russia The distribution of lithological and permafrost facies deposited on the first terrace of Bely Island in the Kara Sea record alternated freezing and thawing cycles. The cryostratigraphy of the terrace, with constraints from data on the taxonomic diversity and habitats of microphytes found in the sediments, allows detailed reconstructions of the permafrost and deposition history associated with Late Pleistocene-Holocene climate and sea level changes. Cryostratigraphy, lithology, сryofacies, microphyte, thermokarst, taberal deposits, sealevel, Kara Sea INTRODUCTION The deposition history of frozen coastal and shelf In the West Arctic, however, the late Quaternary facies and formation of thermokarst in the context of climate and sealevel correlations remain more contro- latest Cenozoic sealevel changes in the Arctic Kara versial (Fig. 1) being interfered with a warming effect Sea area has been much less studied than in the East from the Atlantic water penetrated into the Barents- Arctic shelf. Freezing of shelf sediments in most of the Kara region [Polyakova, 1997]. -
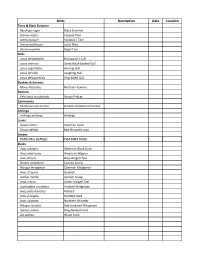
Bird Species Check List
Birds Description Date Location Terns & Black Skimmer Rynchops niger Black Skimmer Sterna caspia Caspian Tern Stema forsteri Forester's Tern Sterna antillarum Least Tern Sterna maxima Royal Tern Gulls Larus philadelphia Bonaparte's Gull Larus marinus Great Black‐backed Gull Larus argentatus Herring Gull Larus atricilla Laughing Gull Larus delawarensis Ring‐billed Gull Boobies & Gannets Morus bassanus Northern Gannet Pelicans Pelecanus occidentalis Brown Pelican Cormorants Phalacrocorax auritus Double‐Crested Cormorant Anhinga Anhinga anhinga Anhinga Loons Gavia immer Common Loon Gavia stellata Red‐throated Loon Grebes PodilymbusPodil ymbus popodicepsdiceps PiPieded‐billbilleded GrebeGrebe Ducks Anas rubripes American Black Duck Anas americana American Wigeon Anas discors Blue‐winged Teal Branta canadensis Canada Goose Mergus merganser Common Merganser Anas strepera Gadwall Aythya marila Greater Scaup Anas crecca Green‐winged Teal Lophodytes cucullatus Hooded Merganser Anas platyrhynchos Mallard Anas fulvigula Mottled Duck Anas clypeata Northern Shoveler Mergus serrator Red‐breasted Merganser Aythya collaris Ring‐Necked Duck Aix sponsa Wood Duck Birds Description Date Location Rails & Northern Jacana Fulica americana American Coot Rallus longirostris Clapper Rail Gallinula chloropus Common Moorhen Rallus elegans King Rail Porzana carolina Sora Long‐legged Waders Nycticorax nycticorax Black‐crowned Night‐Heron Bubulcus ibis Cattle Egret Plegadis falcinellus Glossy Ibis Ardea herodias Great Blue Heron Casmerodius albus Great Egret Butorides -

Radioactivity in the Arctic Seas
IAEA-TECDOC-1075 XA9949696 Radioactivity in the Arctic Seas Report for the International Arctic Seas Assessment Project (IASAP) ffl INTERNATIONAL ATOMIC ENERGY AGENCA / Y / 1JrrziZr^AA 30-16 The originating Section of this publication in the IAEA was: Radiometrics Section International Atomic Energy Agency Marine Environment Laboratory B.P. 800 MC 98012 Monaco Cedex RADIOACTIVITY IN THE ARCTIC SEAS IAEA, VIENNA, 1999 IAEA-TECDOC-1075 ISSN 1011-4289 ©IAEA, 1999 Printe IAEe th AustriAn y i d b a April 1999 FOREWORD From 199 o 1993t e Internationa6th l Atomic Energy Agency's Marine Environment Laboratory (IAEA-MEL s engage IAEA'e wa ) th n di s International Arctic Seas Assessment Project (IASAP whicn i ) h emphasi bees ha sn place criticaa n do l revie f environmentawo l conditions in the Arctic Seas. IAEA-MEe Th L programme, organize framewore th n dIASAi e th f ko P included: (i) an oceanographic and an ecological description of the Arctic Seas; provisioe th (ii )centra a f no l database facilitIASAe th r yfo P programm collectione th r efo , synthesi interpretatiod san datf nmarino n ao e radioactivit Arctie th n yi c Seas; (iii) participation in official expeditions to the Kara Sea organized by the joint Russian- Norwegian Experts Group (1992, 1993 and 1994), the Russian Academy of Sciences (1994), and the Naval Research Laboratory and Norwegian Defence Research Establishment (1995); (iv) assistance wit d n laboratorsiti han u y based radiometric measurement f curreno s t radionuclide concentrations in the Kara Sea; (v) organization of analytical quality assurance intercalibration exercises among the participating laboratories; (vi) computer modellin e potentiath f o g l dispersa f radionuclideo l s released froe mth dumped f assessmeno wast d associatee ean th f o t d radiological consequencee th f o s disposals on local, regional and global scales; (vii) in situ and laboratory based assessment of distribution coefficients (Kd) and concentration factor sArctie (CFth r c)fo environment. -

From the Historiography of the Kamchatka Evens
Journal of Siberian Federal University. Humanities & Social Sciences 5 (2013 6) 713-719 ~ ~ ~ УДК 9-17.094.2/8:636(571.56) From the Historiography of the Kamchatka Evens Antonina G. Koerkova* M.K. Ammosov North-Eastern Federal University in Yakutsk 58 Belinskiy Str., Yakutsk, 677980 Russia Received 11.01.2013, received in revised form 26.03.2013, accepted 30.04.2013 The article explores existing researches on the cultural history of the Evens, who belong to small- numbered indigenous peoples of the North, Siberia, and Far East. The author analyses all the historiographical researches, and classifies them according to the chronology. Keywords: Evens, historiography, small-numbered indigenous peoples of the North, Siberia, and Far East. The work was fulfilled within the framework of the research financed by the Krasnoyarsk Regional Foundation of Research and Technology Development Support and in accordance with the course schedule of Siberian Federal University as assigned by the Ministry of Education and Science of the Russian Federation. From the official sources, the Evens as a life of local peoples. The problem of the research nation are known as the Tungus, Evens, Orochis, history and historiography of the Kamchatka Orochel, Orochons, Lamuts. The most widely Evens is that it has not been studied well. There used names are “evyn”, “evysel” which mean are no integrated monographs dedicated to the “local, living here”. The self-designation of the culture of the Kamchatka Evens. Evens and Ovens are widely spread among the Among the authors who were the first to Evens living in Khabarovsk region, Magadan publish some information on the Kamchatka oblast’, Chukotka Autonomous Okrug. -

Reproduction and Survival in a Declining Population of The
Reproductionand Survivalin a Declining Population of the Southern Dunlin Calidris alpina schinzii Paul Eric J6nsson J6nsson,P.E. 1991.Reproduction and Survival in a DecliningPopulation of theSouthern Dunlin Calidrisalpina schinzii. Wader Study Group Bulletin 61, Supplement: 56-68. Reproductivesuccess and survival was studied in a declining population of theSouthern Dunlin in SW Sk,Sne,SSweden, in 1981-1986.Hatching success was low; on average only 30% of clutches hatched, dueto intense nest-predation from mainly Crows, Foxes and mustelids. Some nests (8%) were also lostby tramplingfrom grazing animals. Within a season,there was no significantdifference in hatchingsuccess between early and late clutches, but the overall annual hatching success was higher in yearswith an early onset of breeding.On average,the Dunlins yearly produced 0.3 - 0.4 hatched young/adult.Fledging success was estimated at36% and survival from fledging tothe age of one year at56%. Adult survival, estimated from rerum rates, averaged 89% in males and 77% in females (83% of thesexes combined). The sex-relateddifference in return-ratesbelieved to reflecta loweractuai survivalrate in females, assuggested bythe male-skewed sex-ratio inthe population. The average life expectancyfor an adult Dunlin is estimated at5.4 years and the mean longevity at7.4 years. Survival tendedto decrease with increasing age (up to 5 yearsafter ringing). The oldest bird recorded in the studywas 17 yearsold. The observed reproductive rate was found to be insufficientto maintaina stablepopulation and unless nest-predation issignificantly reduced, the Dunlin will become extinct in SW Sl•ne within20 to 30 years. PaulE. J6nsson,Dept. of Ecology, Ecology Building, University of Lund, 223 62 Lund, Sweden. -

Breeding Schedule and Primary Moult in Dunlins <I>Calidris Alpina</I> Of
29 Breeding scheduleand primary moult in Dunlins Calidris alpina of the Far East Panel S. Tomkoich Tomkovich,P.S. 1998. Breedingschedule and primarymoult in Dunlins Calidris alpina of the Far East. WaderStudy Group Bull. 85:29-34 On Chukotka,the breedinggrounds of the subspeciessakhalina, the primarymoult of Dunlin Calidrisalpina startsin mid-June,at the beginningof theincubation period, and birds migrate southward from thebeginning of August,when the moultis almostcomplete. On Kamchatka(kistchinski) and Sakhalin(actites), where breeding takes place earlier, primary moult startsin July,in the chick rearing period,or evenlater. Thesedifferences between northern and southern populations are similarto thosefound previously in Alaska (Holmes1971). The timing of theseevents is not strictlyrelated to latitude; it is thoughtthat, with localvariations, latitudinal differencesin datesof breedingand primary moult are brought about through subspecific characteristics. Knowledge of subspecific breedingschedules and primary moult periods can be usedtogether with biometricsto assignFar EasternDunlins to different populationsin the post-breedingperiod. P S. Tomkovich,Zoological Museum, Moscow Lomonosov State University,Bolshaya Nikitskaya Street 6, Moscow103009, Russia. INTRODUCTION Far East are comparedin this study. Two new subspeciesof Dunlin from the Far East, Calidris alpina kistchinski(Tornkovich 1986) and C.a. actites(Nechaev STUDY AREAS AND METHODS & Tornkovich1987, 1988), were describedrecently, in The main data were collectedduring expeditions -

Translitterering Och Alternativa Geografiska Namnformer
TRANSLITTERERING OCH ALTERNATIVA GEOGRAFISKA NAMNFORMER Version XX, 27 juli 2015, Stefan Nordblom 1 FÖRORD För många utländska egennamn, i första hand personnamn och geografiska namn, finns det på svenska väl etablerade namnformer. Om det inte finns någon sådan kan utländska egennamn dock vålla bekymmer vid översättning till svenska. Föreliggande material är tänkt att vara till hjälp i sådana situationer och tar upp fall av translitterering1 och transkribering2 samt exonymer3 . Problemen uppstår främst på grund av att olika språk har olika system för translitterering och transkribering från ett visst språk och på grund av att orter kan ha olika namn på olika utländska språk. Eftersom vi oftast översätter från engelska och franska innehåller sammanställningen även translittereringar och exonymer på engelska och franska (samt tyska). Man kan alltså i detta material göra en sökning på sådana namnformer och komma fram till den svenska namnformen. Om man t.ex. i en engelsk text träffar på det geografiska namnet Constance kan man söka på det namnet här och då få reda på att staden (i detta fall på tyska och) på svenska kallas Konstanz. Den efterföljande sammanställningen bygger i huvudsak på följande källor: Institutet för de inhemska språken (FI): bl.a. skriften Svenska ortnamn i Finland - http://kaino.kotus.fi/svenskaortnamn/ Iate (EU-institutionernas termbank) Nationalencyklopedin Nationalencyklopedins kartor Interinstitutionella publikationshandboken - http://publications.europa.eu/code/sv/sv-000100.htm Språkbruk (Tidskrift utgiven av Svenska språkbyrån i Helsingfors) Språkrådet© (1996). Publikation med rekommendationer i term- och språkfrågor som utarbetas av rådets svenska översättningsenhet i samråd med övriga EU-institutioner. TT-språket - info.tt.se/tt-spraket/ I de fall uppgifterna i dessa källor inte överensstämmer med varandra har det i enskilda fall varit nödvändigt att väga, välja och sammanjämka namnförslagen, varvid rimlig symmetri har eftersträvats. -

Social Transition in the North, Vol. 1, No. 4, May 1993
\ / ' . I, , Social Transition.in thb North ' \ / 1 \i 1 I '\ \ I /? ,- - \ I 1 . Volume 1, Number 4 \ I 1 1 I Ethnographic l$ummary: The Chuko tka Region J I / 1 , , ~lexdderI. Pika, Lydia P. Terentyeva and Dmitry D. ~dgo~avlensly Ethnographic Summary: The Chukotka Region Alexander I. Pika, Lydia P. Terentyeva and Dmitry D. Bogoyavlensky May, 1993 National Economic Forecasting Institute Russian Academy of Sciences Demography & Human Ecology Center Ethnic Demography Laboratory This material is based upon work supported by the National Science Foundation under Grant No. DPP-9213l37. Any opinions, findings, and conclusions or recammendations expressed in this material are those of the author@) and do not ncccssarily reflect the vim of the National Science Foundation. THE CHUKOTKA REGION Table of Contents Page: I . Geography. History and Ethnography of Southeastern Chukotka ............... 1 I.A. Natural and Geographic Conditions ............................. 1 I.A.1.Climate ............................................ 1 I.A.2. Vegetation .........................................3 I.A.3.Fauna ............................................. 3 I1. Ethnohistorical Overview of the Region ................................ 4 IIA Chukchi-Russian Relations in the 17th Century .................... 9 1I.B. The Whaling Period and Increased American Influence in Chukotka ... 13 II.C. Soviets and Socialism in Chukotka ............................ 21 I11 . Traditional Culture and Social Organization of the Chukchis and Eskimos ..... 29 1II.A. Dwelling .............................................. -

K. Fujita1, E. E. Dretzka2 and A. Grantz3 This Report Is Preliminary and Has Not Been Reviewed for Conformity with U. S. Geologi
UNITED STATES DEPARTMENT OF THE INTERIOR GEOLOGICAL SURVEY BIBLIOGRAPHY OF NORTHEAST SIBERIAN GEOLOGY AND GEOPHYSICS (SECOND EDITION) K. Fujita1, E. E. Dretzka2 and A. Grantz3 OPEN FILE REPORT 82-616 This report is preliminary and has not been reviewed for conformity with U. S. Geological Survey editorial standards. Menlo Park, California July 1982 1. Department of Geology, Michigan State University, East Lansing MI 48824 2. Department of Geological Sciences, Northwestern University, Evanston IL 60201; now at: Department of Computer Sciences, Stanford University, Stanford CA 94305. 3. U. S. Geological Survey, Menlo Park, CA 94025 INTRODUCTION This bibliography is a compendium of literature available in English, either in its original form or in translation, on the geology and geophysics of northeast Siberia and adjacent seas and shelves. It is an expanded version of Fujita and Dretzka (1978-) and has been updated to include publications released through early 1982. The area! coverage extends from the edge of the Siberian platform (just east of the Lena River) on the east to the U. S. - Russia Convention Line of 1867 on the west. The East Siberian and Chukchi Seas are included as the northern limit while the Sea if Okhotsk and Kamchatka represent the southern limits. Sakhalin has been excluded and some, but not all, references to the Kuril Islands have been included. A sketch map of the area is shown in figure 1. It is hoped that this listing is nearly exhaustive for works on this area with some exceptions. Neither the Paleontological Journal nor Petroleum Geology have been indexed in this edition and, in addition, articles on Recent seismicity and volcanic activity in the Kuril-Kamchatka arc have been omitted. -

Peeps and Related Sandpipers Peeps Are a Group of Diminutive Sandpipers That Are Notoriously Hard to Tell Apart
Peeps and Related Sandpipers Peeps are a group of diminutive sandpipers that are notoriously hard to tell apart. They belong to a subfamily of subarctic and arctic nesting sandpipers known as the Calidridinae (in the sandpiper family, Scolopacidae). During their migrations, when most residents of North America have the opportunity to watch them, mixed flocks of calidridine sandpipers scurry about on mudflats, feeding at the edge of the retreating tide, or swarm aloft, twisting and turning like a dense school of fish. These traits, in a group of birds that look so much alike to start with, give bird watchers nightmares. Fortunately for Alaskans and visitors to our state, Alaska is an excellent location to view and identify calidridine sandpipers. The early summer breeding season is the easiest time of the year to distinguish the various species, not only because they are in breeding plumage and are more approachable than at other times of the year, but also because each species performs a characteristic courtship display with unique vocalizations. For the avid birder, Alaska has the additional attraction of being one of the best places in North America to view exotic Eurasian species. General description: Three peeps are abundant summer residents and breeders in Alaska—the least, semipalmated, and western sandpipers (Calidris minutilla, C. pusilla, and C. mauri) [all lists in order by size]. Another four species from Eurasia may also be seen—the little, rufous-necked, Temminck's, and long-toed stints (“stint” is the British equivalent for peep) (C. minuta, C. ruficollis, C. temminckii, C. subminuta). These seven species range from 5 to 6½ inches (15-17 cm) in length, and weigh from 2/3 to 1½ ounces (17-33 g).