The Dinosaurs of the “Continental Intercalaire” of the Central Sahara by A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

BGS Report, Single Column Layout
1 The Cretaceous Continental Intercalaire in central Algeria: subsurface evidence for a fluvial to aeolian 2 transition and implications for the onset of aridity on the Saharan Platform 3 A.J. Newella*, G.A. Kirbyb, J.P.R. Sorensena, A.E. Milodowskib 4 a British Geological Survey, Maclean Building, Wallingford, OX10 8BB, UK, email: [email protected] 5 b British Geological Survey, Nicker Hill, Keyworth, Nottingham, NG12 5GG, UK 6 * Corresponding author 7 Abstract 8 The Lower Cretaceous Continental Intercalaire of North Africa is a terrestrial to shallow marine 9 continental wedge deposited along the southern shoreline of the Neotethys Ocean. Today it has a wide 10 distribution across the northern Sahara where it has enormous socio-economic importance as a major 11 freshwater aquifer. During the Early Cretaceous major north-south trending basement structures were 12 reactivated in response to renewed Atlantic rifting and in Algeria, faults along the El Biod-Hassi 13 Messaourd Ridge appear to have been particularly important in controlling thickness patterns of the 14 Lower Cretaceous Continental Intercalaire. Subsurface data from the Krechba gas field in Central Algeria 15 shows that the Lower Cretaceous stratigraphy is subdivided into two clear parts. The lower part (here 16 termed the In Salah Formation) is a 200 m thick succession of alluvial deposits with large meandering 17 channels, clearly shown in 3D seismic, and waterlogged flood basins indicated by lignites and gleyed, 18 pedogenic mudstones. The overlying Krechba Formation is a 500 m thick succession of quartz- 19 dominated sands and sandstones whose microstructure indicates an aeolian origin, confirming earlier 20 observations from outcrop. -

Biology Assessment Plan Spring 2019
Biological Sciences Department 1 Biology Assessment Plan Spring 2019 Task: Revise the Biology Program Assessment plans with the goal of developing a sustainable continuous improvement plan. In order to revise the program assessment plan, we have been asked by the university assessment committee to revise our Students Learning Outcomes (SLOs) and Program Learning Outcomes (PLOs). Proposed revisions Approach: A large community of biology educators have converged on a set of core biological concepts with five core concepts that all biology majors should master by graduation, namely 1) evolution; 2) structure and function; 3) information flow, exchange, and storage; 4) pathways and transformations of energy and matter; and (5) systems (Vision and Change, AAAS, 2011). Aligning our student learning and program goals with Vision and Change (V&C) provides many advantages. For example, the V&C community has recently published a programmatic assessment to measure student understanding of vision and change core concepts across general biology programs (Couch et al. 2019). They have also carefully outlined student learning conceptual elements (see Appendix A). Using the proposed assessment will allow us to compare our student learning profiles to those of similar institutions across the country. Revised Student Learning Objectives SLO 1. Students will demonstrate an understanding of core concepts spanning scales from molecules to ecosystems, by analyzing biological scenarios and data from scientific studies. Students will correctly identify and explain the core biological concepts involved relative to: biological evolution, structure and function, information flow, exchange, and storage, the pathways and transformations of energy and matter, and biological systems. More detailed statements of the conceptual elements students need to master are presented in appendix A. -

Titanosauriform Teeth from the Cretaceous of Japan
“main” — 2011/2/10 — 15:59 — page 247 — #1 Anais da Academia Brasileira de Ciências (2011) 83(1): 247-265 (Annals of the Brazilian Academy of Sciences) Printed version ISSN 0001-3765 / Online version ISSN 1678-2690 www.scielo.br/aabc Titanosauriform teeth from the Cretaceous of Japan HARUO SAEGUSA1 and YUKIMITSU TOMIDA2 1Museum of Nature and Human Activities, Hyogo, Yayoigaoka 6, Sanda, 669-1546, Japan 2National Museum of Nature and Science, 3-23-1 Hyakunin-cho, Shinjuku-ku, Tokyo 169-0073, Japan Manuscript received on October 25, 2010; accepted for publication on January 7, 2011 ABSTRACT Sauropod teeth from six localities in Japan were reexamined. Basal titanosauriforms were present in Japan during the Early Cretaceous before Aptian, and there is the possibility that the Brachiosauridae may have been included. Basal titanosauriforms with peg-like teeth were present during the “mid” Cretaceous, while the Titanosauria with peg-like teeth was present during the middle of Late Cretaceous. Recent excavations of Cretaceous sauropods in Asia showed that multiple lineages of sauropods lived throughout the Cretaceous in Asia. Japanese fossil records of sauropods are conformable with this hypothesis. Key words: Sauropod, Titanosauriforms, tooth, Cretaceous, Japan. INTRODUCTION humerus from the Upper Cretaceous Miyako Group at Moshi, Iwaizumi Town, Iwate Pref. (Hasegawa et al. Although more than twenty four dinosaur fossil local- 1991), all other localities provided fossil teeth (Tomida ities have been known in Japan (Azuma and Tomida et al. 2001, Tomida and Tsumura 2006, Saegusa et al. 1998, Kobayashi et al. 2006, Saegusa et al. 2008, Ohara 2008, Azuma and Shibata 2010). -

Transfer of the Type Species of the Genus Themobacteroides to the Genus Themoanaerobacter As Themoanaerobacter Acetoethylicus (Ben-Bassat and Zeikus 1981) Comb
INTERNATIONAL JOURNAL OF SYSTEMATICBACTERIOLOGY, Oct. 1993, p. 857-859 Vol. 43, No. 4 0020-7713/93/040857-03$02.00/0 Copyright 0 1993, International Union of Microbiological Societies Transfer of the Type Species of the Genus Themobacteroides to the Genus Themoanaerobacter as Themoanaerobacter acetoethylicus (Ben-Bassat and Zeikus 1981) comb. nov., Description of Coprothemobacter gen. nov., and Reclassification of Themobacteroides proteolyticus as Coprothennobacter proteolyticus (Ollivier et al. 1985) comb. nov. FRED A. RAINEY AND ERKO STACKEBRANDT* DSM-Deutsche Sammlung von Mikroolganisrnen und Zellkulturen, Mascheroder Weg lb, 38124 Braunschweig, Germany Phylogenetic and phenotypic evidence demonstrates the taxonomic heterogeneity of the genus Thennobac- teroides and indicates a close relationship between Thennobacteroides acetoethylicus and members of the genus Thennoanaerobacter. Since T. acetoethylicus is the type species of Thennobacteroides, its removal invalidates the genus. As a consequence, the remaining species Thennobacteroides proteolyticus is proposed as the type species of the new genus Coprothennobacter gen. nov., as Coprothennobacter proteolyticus comb. nov. Recent phylogenetic studies (8, 9) of anaerobic thermo- hybridization studies with all members of Thermoanaero- philic species demonstrated that the majority of strains fall bacter (6, 10) despite the fact that, like Thermoanaerobacter within the phylogenetic confines of the Clostridium-Bacillus species, Thermobacteroides acetoethylicus is an anaerobic, subphylum of gram-positive bacteria. In contrast to the thermophilic, glycolytic bacterium capable of growth above phylog enet ically coherent genera Thermoana erobac ter (6) 70°C that has been isolated from geothermal environments. and Thermoanaerobacterium (6), members of Thermobac- The reclassifications of the recent study of Lee et al. (6) teroides (1,7) belonged to phylogenetically very diverse taxa increased the numbers of species of Thermoanaerobacter to (8). -

Re-Description of the Sauropod Dinosaur Amanzia (“Ornithopsis
Schwarz et al. Swiss J Geosci (2020) 113:2 https://doi.org/10.1186/s00015-020-00355-5 Swiss Journal of Geosciences ORIGINAL PAPER Open Access Re-description of the sauropod dinosaur Amanzia (“Ornithopsis/Cetiosauriscus”) greppini n. gen. and other vertebrate remains from the Kimmeridgian (Late Jurassic) Reuchenette Formation of Moutier, Switzerland Daniela Schwarz1* , Philip D. Mannion2 , Oliver Wings3 and Christian A. Meyer4 Abstract Dinosaur remains were discovered in the 1860’s in the Kimmeridgian (Late Jurassic) Reuchenette Formation of Moutier, northwestern Switzerland. In the 1920’s, these were identifed as a new species of sauropod, Ornithopsis greppini, before being reclassifed as a species of Cetiosauriscus (C. greppini), otherwise known from the type species (C. stewarti) from the late Middle Jurassic (Callovian) of the UK. The syntype of “C. greppini” consists of skeletal elements from all body regions, and at least four individuals of diferent sizes can be distinguished. Here we fully re-describe this material, and re-evaluate its taxonomy and systematic placement. The Moutier locality also yielded a theropod tooth, and fragmen- tary cranial and vertebral remains of a crocodylomorph, also re-described here. “C.” greppini is a small-sized (not more than 10 m long) non-neosauropod eusauropod. Cetiosauriscus stewarti and “C.” greppini difer from each other in: (1) size; (2) the neural spine morphology and diapophyseal laminae of the anterior caudal vertebrae; (3) the length-to-height proportion in the middle caudal vertebrae; (4) the presence or absence of ridges and crests on the middle caudal cen- tra; and (5) the shape and proportions of the coracoid, humerus, and femur. -

The Continental Intercalaire Aquifer at the Kébili Geothermal Field, Southern Tunisia
Proceedings World Geothermal Congress 2005 Antalya, Turkey, 24-29 April 2005 The Continental Intercalaire Aquifer at the Kébili Geothermal Field, Southern Tunisia Aissa Agoun Regional Commissariat for Agricultural Development Water Resources Departement, C.R.D.A Kébili, Kébili 4200, TUNISIA [email protected] Keywords: CI, Kébili, wells, artesian wellheads like valves and monitoring points. Also drawdown of the water level has been observed due to the ABSTRACT increasing water demands. Radiocarbon analysis has shown that radiocarbon is present at between 2 and 10 pmc which The C.I. "Continental Intercalaire" aquifer is an extensive leads to the conclusion that the water is recharged during horizontal sandstone reservoir (Neocomien: Lower the late Pleistocene 25,000 years B.P, corresponding to the Cretaceous: Purbecko-Wealdien). The CI is one of the last glaciation (Edmonds et al., 1997). Water is fossil and largest aquifers in the world covering more than 1 million 2 without actual recharge, so to preserve the trans-borderers km in Tunisia, Algeria and Libya. This aquifer covers reservoir, more research must be carried out on the aquifer. 80,000 km2 in Tunisia Full collaboration with Algerian and Libyan organizations In Kébili field in southern Tunisia, the geothermal water is is necessary in order to achieve economical and sustainable about 25 to 50 thousand years old and of sulphate chlorite future production and regional development. Long term type. The depth of the reservoir ranges from 1000 to 2800 monitoring of pressure temperature and salinity at the m. wellhead for each production well should be carried out. Any increase of drawdown during the next 20 years caused The piézométric level is about 200 meters above the by increased production will lead to environmental changes. -

A New Camarasaurid Sauropod Opisthocoelicaudia Skarzynskii Gen
MAGDALENA BORSUK-BIALYNICKA A NEW CAMARASAURID SAUROPOD OPISTHOCOELICAUDIA SKARZYNSKII GEN. N., SP. N. FROM THE UPPER CRETACEOUS OF MONGOLIA (plates 1-14) Abstract. - An almost complete postcranial skeleton lacking cervicals of Opisthocoelicaudia skarzynskii gen. n., sp. n. (Sauropoda, Camarasauridae) from the Upper Cretaceous Nemegt Formation, Gobi Desert , is described and figured. The reconstruction of the muscle system and sternum as well as the restoration of the whole animal is made. It is shown that Opisthocoelicaudia was a straight backed sauropod with the tail carried in a horizontal position. The neck is supposed to have been of medium length (about 5 m) and was carried low. The possibility of habitual assuming a tirpodal position is suggested by the opisthocoelous structure of the ant erior caudals. The importance of some osteologic features of sauropods for the understanding of their attitudes as well as for the systematics is discussed. It is argued that the length of neural spines depends on both the curvature of the back-bone and the length of the neck and tail in sauropods. Forked neural spines are indicative ot the habitual lowering of the neck, or even of the low carrying of the neck, if the anterior dorsals lack traces of the nuchal ligament insertion. Some titanosaurid characters of Opisthocoelicaudia are regarded as progressive ones in sauropods, whereas its camarasaurid features seem to indicate a true relationship in spite of their highly behavioural character. CONTENTS Page Introduction. 6 Description ... 8 Vertebral column 9 Thoracic ribs . 18 Sternum . 19 Pectoral girdle 22 Fore limbs . 24 Pelvic girdle . 32 Hind limbs . -
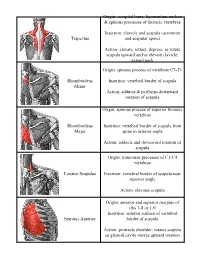
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

Dinosaur Species List E to M
Dinosaur Species List E to M E F G • Echinodon becklesii • Fabrosaurus australis • Gallimimus bullatus • Edmarka rex • Frenguellisaurus • Garudimimus brevipes • Edmontonia longiceps ischigualastensis • Gasosaurus constructus • Edmontonia rugosidens • Fulengia youngi • Gasparinisaura • Edmontosaurus annectens • Fulgurotherium australe cincosaltensis • Edmontosaurus regalis • Genusaurus sisteronis • Edmontosaurus • Genyodectes serus saskatchewanensis • Geranosaurus atavus • Einiosaurus procurvicornis • Gigantosaurus africanus • Elaphrosaurus bambergi • Giganotosaurus carolinii • Elaphrosaurus gautieri • Gigantosaurus dixeyi • Elaphrosaurus iguidiensis • Gigantosaurus megalonyx • Elmisaurus elegans • Gigantosaurus robustus • Elmisaurus rarus • Gigantoscelus • Elopteryx nopcsai molengraaffi • Elosaurus parvus • Gilmoreosaurus • Emausaurus ernsti mongoliensis • Embasaurus minax • Giraffotitan altithorax • Enigmosaurus • Gongbusaurus shiyii mongoliensis • Gongbusaurus • Eoceratops canadensis wucaiwanensis • Eoraptor lunensis • Gorgosaurus lancensis • Epachthosaurus sciuttoi • Gorgosaurus lancinator • Epanterias amplexus • Gorgosaurus libratus • Erectopus sauvagei • "Gorgosaurus" novojilovi • Erectopus superbus • Gorgosaurus sternbergi • Erlikosaurus andrewsi • Goyocephale lattimorei • Eucamerotus foxi • Gravitholus albertae • Eucercosaurus • Gresslyosaurus ingens tanyspondylus • Gresslyosaurus robustus • Eucnemesaurus fortis • Gresslyosaurus torgeri • Euhelopus zdanskyi • Gryponyx africanus • Euoplocephalus tutus • Gryponyx taylori • Euronychodon -

Influence of Basement Heterogeneity on the Architecture of Low Subsidence Rate Paleozoic
Solid Earth Discuss., https://doi.org/10.5194/se-2018-50 Manuscript under review for journal Solid Earth Discussion started: 27 June 2018 c Author(s) 2018. CC BY 4.0 License. 1 Influence of basement heterogeneity on the architecture of low subsidence rate Paleozoic 2 intracratonic basins (Ahnet and Mouydir basins, Central Sahara) 3 Paul Perron1, Michel Guiraud1, Emmanuelle Vennin1, Isabelle Moretti2, Éric Portier3, Laetitia 4 Le Pourhiet4, Moussa Konaté5 5 1Université de Bourgogne Franche-Comté, Centre des Sciences de la Terre, UMR CNRS 6 6282 Biogéosciences, 6 Bd Gabriel, 21000 Dijon, France. 7 2ENGIE, Département Exploration & Production, 1, place Samuel de Champlain, Faubourg 8 de l'Arche, 92930 Paris La Défense, France. 9 3NEPTUNE Energy International S.A., 9-11 Allée de l'Arche – Tour EGEE – 92400 10 Courbevoie, France. 11 4Sorbonne Université, CNRS-INSU, Institut des Sciences de la Terre Paris, ISTeP UMR 12 7193, F-75005 Paris, France. 13 5Département de Géologie, Université Abdou Moumouni de Niamey, BP :10662, Niamey, 14 Niger. 15 Corresponding author: [email protected], [email protected] 16 Abstract 17 The Paleozoic intracratonic North African Platform is characterized by an association of 18 arches (ridges, domes, swells or paleo-highs) and low subsidence rate syncline basins of 19 different wavelengths (75–620 km). The structural framework of the platform results from the 20 accretion of Archean and Proterozoic terranes during the Pan-African orogeny (750–580 Ma). 21 The Ahnet and Mouydir basins are successively delimited from east to west by the Amguid El 22 Biod, Arak-Foum Belrem, and Azzel Matti arches, bounded by inherited Precambrian sub- 23 vertical fault systems which were repeatedly reactivated or inverted during the Paleozoic. -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks. -

The Dinosaurs of North America
FEOM THE SIXTEENTH ANNUAL KEPOKT OF THE U. S, GEOLOGICAL SURVEY THE DINOSAURS OF NORTH AMERICA OTHNIEL CHARLES MARSH TALE UNIVERSITY WASHINGTON 1896 ^33/^, I/BRAKt 4 ,\ . THE DINOSAURS OF NORTH AMERICA. BY OTHNIEL CHARLES MARSH. 133 CONTENTS. Pajje. Introduction 143 Part I. —Triassic dinosaurs 146 Theropoda 146 Anchisaurida? 147 Anchisaurus 147 The skull 148 The fore limbs 149 The hind limbs 149 Anchisaurus solus 149 Amniosaurus rTT 150 Eestoration of Anchisaurus 150 Dinosaurian footprints 151 Distribution of Triassic dinosaurs 152 Part II. —Jurassic dinosaurs 152 Theropoda : 153 Hallopus 153 Fore and hind limbs 154 Coelurus _ 155 The vertebra:- 155 The hind limbs 156 Ceratosaurus 156 The skull 157 The brain 159 The lower iaws 159 The vertebra: 159 The scapular arch 160 The pelvic arch 160 The metatarsals 162 Eestoration of Ceratosaurus 163 Allosaurus 163 European Theropoda 163 Sauropoda 164 Atlantosaurus beds 164 Families of Sauropoda 165 Atlantosauridie 166 Atlantosaurus 166 Apatosaurus 166 The sacral cavity 166 The vertebra- 167 Brontosaurus 168 The scapular arch 168 The cervical vertebra.- 169 The dorsal vertebree 169 The sacrum 170 The caudal vertebra- 171 The pelric arch 172 The fore limbs 173 The hind limbs 173 135 136 CONTENTS. Part II. —Jurassic dinosaurs—Continued. Page. Sauropoda—Continued. Atlantosaurida? —Continued. Restoration of Brontosaurus 173 Barnsaurus 174 Diplodoeida? 175 Diplodocus 175 The skull 175 The brain 178 The lower jaws 178 The teeth 179 The vertebra; 180 The sternal bones 180 The pelvic girdle 180 Size and habits 180 Morosaurida? 181 Morosaurus 181 The skull 181 The vertebra? 181 The fore limbs 182 The pelvis 182 The hind limbs 183 Pleuroccelida? : 183 Pleurocoelus 183 The skull 183 The vertebras 183 Distribution of the Sauropoda 185 Comparison with European forms 185 Predentata ».