Immobilization of P. Digitatum and Bioconversion of Limonene to Alpha-Terpineol. Qiang Tan Louisiana State University and Agricultural & Mechanical College
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Retention Indices for Frequently Reported Compounds of Plant Essential Oils
Retention Indices for Frequently Reported Compounds of Plant Essential Oils V. I. Babushok,a) P. J. Linstrom, and I. G. Zenkevichb) National Institute of Standards and Technology, Gaithersburg, Maryland 20899, USA (Received 1 August 2011; accepted 27 September 2011; published online 29 November 2011) Gas chromatographic retention indices were evaluated for 505 frequently reported plant essential oil components using a large retention index database. Retention data are presented for three types of commonly used stationary phases: dimethyl silicone (nonpolar), dimethyl sili- cone with 5% phenyl groups (slightly polar), and polyethylene glycol (polar) stationary phases. The evaluations are based on the treatment of multiple measurements with the number of data records ranging from about 5 to 800 per compound. Data analysis was limited to temperature programmed conditions. The data reported include the average and median values of retention index with standard deviations and confidence intervals. VC 2011 by the U.S. Secretary of Commerce on behalf of the United States. All rights reserved. [doi:10.1063/1.3653552] Key words: essential oils; gas chromatography; Kova´ts indices; linear indices; retention indices; identification; flavor; olfaction. CONTENTS 1. Introduction The practical applications of plant essential oils are very 1. Introduction................................ 1 diverse. They are used for the production of food, drugs, per- fumes, aromatherapy, and many other applications.1–4 The 2. Retention Indices ........................... 2 need for identification of essential oil components ranges 3. Retention Data Presentation and Discussion . 2 from product quality control to basic research. The identifi- 4. Summary.................................. 45 cation of unknown compounds remains a complex problem, in spite of great progress made in analytical techniques over 5. -

Grapefruit Organic Botanical Name: Citrus X Paradisi Origin: Isreal
GC/MS BATCH NUMBER: GK0102 ESSENTIAL OIL: GRAPEFRUIT ORGANIC BOTANICAL NAME: CITRUS X PARADISI ORIGIN: ISREAL KEY CONSTITUENTS PRESENT IN THIS BATCH OF % GRAPEFRUIT ORGANIC OIL LIMONENE 92.9 MYRCENE 1.8 Comments from Robert Tisserand: Exquisite pink Grapefruit odor quality. Does not quite meet the full ISO standards, but this looks like a genuine oil. 510 2nd St S. Twin Falls, ID 83301 * 800-917-6577 * planttherapy.com facebook.com/PlantTherapy * planttherapy.com/blog Date : March 05, 2018 CERTIFICATE OF ANALYSIS - GC PROFILING SAMPLE IDENTIFICATION Internal code : 18C01-PTH2-1-CC Customer identification : Grapefruit Organic - Israel - GK0102191R Type : Essential oil Source : Citrus x paradisi cv. Yellow Customer : Plant Therapy ANALYSIS Method: PC-PA-014-17J19 - Analysis of the composition of an essential oil, or other volatile liquid, by FAST GC-FID (in French); identifications validated by GC-MS. Analyst : Sylvain Mercier, M. Sc., Chimiste Analysis date : March 05, 2018 Checked and approved by : Alexis St-Gelais, M. Sc., chimiste 2013-174 Note: This report may not be published, including online, without the written consent from Laboratoire PhytoChemia. This report is digitally signed, it is only considered valid if the digital signature is intact. Page 1/9 Essential oil, Citrus x paradisi cv. Yellow Report prepared for Internal code: 18C01-PTH2-1-CC Grapefruit Organic - Israel - GK0102191R Plant Therapy PHYSICOCHEMICAL DATA Physical aspect: Bright yellow liquid Refractive index: 1.4745 ± 0.0003 (20 °C) CONCLUSION No adulterant, contaminant -

An Artifact in a Synthetic Pine Oil
RESEARCH NOTE J. Ess. Oil Res., 3, 41-42 (Jan/Feb 1991) An Artifact in a Synthetic Pine Oil Duane F. Zinkel USDA Forest Service, Forest Products Laboratory* One Gifford Pinchot Drive Madison, WI 53705-2398 ABSTRACT: The isopropyl ether of a-terpineol was identified as an artifact in the synthetic pine oil produced when isopropyl alcohol was used as the emulsifier. KEY WORD INDEX: Synthetic pine oil, a-terpineol isopropyl ether, terpinen-4-ol isopropyl ether, turpentine. INTRODUCTION: The manufacture of synthetic pine oil is the primary use for turpentine. The synthesis involves the acid-catalyzed hydration of a-pinene at the in terface ofan emulsion of pinene/mineral acid (1). Various emulsifiers have been used, one of which is isopropyl alcohol. Our gas chromatographic examination of a commercial distilled pine oil, produced using the isopropyl alcohol emulsifier, revealed the presence of 4-5% of a higher boiling component product not present originally in the turpentine. EXPERIMENTAL: NMR spectra were obtained at 310 K with a Bruker WM250 (250 MHz proton and 62.9 MHz carbon) FT spectrometer controlled by an Aspect 2000A minicomputer; DEPT spectra were obtained with a standard Bruker program. Gas chromatography was done with a Hewlett Packard 5880 gas chromatograph (FID) and fused-silica columns: a DB-1 (a methyl silicone) column from J & W Scientific (Folsom, CA), 15m x 0.25mmi.d. witha 0.1-µmfilmoperatedat60°Cand a Carbowaxcolumn,30m x 0.25mm with a 0.25-µm film temperature programmed from 60°C to 225°C at 8°C/min. isopropyl etherwas isolated by liquid chromatography. -

(19) United States (12) Patent Application Publication (10) Pub
US 20060081822A1 (19) United States (12) Patent Application Publication (10) Pub. N0.: US 2006/0081822 A1 Koetzle (43) Pub. Date: Apr. 20, 2006 (54) METHOD TO INCREASE FLASH POINTS OF (52) US. Cl. ............................................................ .. 252/601 FLAMMABLE SOLVENTS (76) Inventor: A. Richard Koetzle, Rochester, NY (57) ABSTRACT (Us) Correspondence AddreSSI The present invention relates to a method to decrease the A- RICHARD KOETZLE ?ammability of normally ?ammable alcohols and solvents. 134 STONECLIFF DRIVE The additive is Alpha Terpineol, Which Will increase the ROCHESTER’ NY 14616 (Us) ?ash point of ?ammable alcohols or solvents, by blending the Terpineol into the ?ammable solvent or alcohol. Solvents (21) Appl' NO" 10/968’441 such as acetone, methanol, ethylacetate, ethanol and Xylene, (22) Filed, Oct 20 2004 to name a feW, increases ?ash points by 50° C. to 60° C., by i ’ addition of 12-14% terpineol. The said solvent can then be Publication Classi?cation blended With other organic solvents to produce performance solvents, such as paint strippers With ?ash points greater (51) Int. Cl. than 1400 F. and meet Federal and state Volatile Organic C 09K 21/00 (2006.01) Compound regulations. US 2006/0081822 A1 Apr. 20, 2006 METHOD TO INCREASE FLASH POINTS OF [0008] The organic solvent or combination of solvents can FLAMMABLE SOLVENTS comprise up to 99 Weight percent of the composition in total, and may be the combination of tWo or more different types BACKGROUND OF THE INVENTION of organic solvents. A typical combination may comprise; [0001] Many industrial processing cleaning compositions [0009] 1.0 to 99 Weight percent organic solvent. -

Neuroprotective Potential of Limonene and Limonene Containing Natural Products
molecules Review Neuroprotective Potential of Limonene and Limonene Containing Natural Products Lujain Bader Eddin 1 , Niraj Kumar Jha 2 , M. F. Nagoor Meeran 1 , Kavindra Kumar Kesari 3,4 , Rami Beiram 1 and Shreesh Ojha 1,* 1 Department of Pharmacology and Therapeutics, College of Medicine and Health Sciences, United Arab Emirates University, P.O. Box 17666, Al Ain 17666, United Arab Emirates; [email protected] (L.B.E.); [email protected] (M.F.N.M.); [email protected] (R.B.) 2 Department of Biotechnology, School of Engineering & Technology, Sharda University, Greater Noida, Uttar Pradesh 201310, India; [email protected] 3 Department of Applied Physics, School of Science, Aalto University, 00076 Espoo, Finland; kavindra.kesari@aalto.fi 4 Department of Bioproducts and Biosystems, School of Chemical Engineering, Aalto University, 00076 Espoo, Finland * Correspondence: [email protected] Abstract: Limonene is a monoterpene confined to the family of Rutaceae, showing several biological properties such as antioxidant, anti-inflammatory, anticancer, antinociceptive and gastroprotective characteristics. Recently, there is notable interest in investigating the pharmacological effects of limonene in various chronic diseases due to its mitigating effect on oxidative stress and inflam- mation and regulating apoptotic cell death. There are several available studies demonstrating the neuroprotective role of limonene in neurodegenerative diseases, including Alzheimer’s disease, Citation: Eddin, L.B.; Jha, N.K.; multiple sclerosis, epilepsy, anxiety, and stroke. The high abundance of limonene in nature, its Meeran, M.F.N.; Kesari, K.K.; Beiram, safety profile, and various mechanisms of action make this monoterpene a favorable molecule to R.; Ojha, S. -

Laboratory Study of Polychlorinated Biphenyl (PCB) Contamination and Mitigation in Buildings Part 4. Evaluation of the Activate
EPA/600/R-11/156C November 2012 Laboratory Study of Polychlorinated Biphenyl (PCB) Contamination and Mitigation in Buildings Part 4. Evaluation of the Activated Metal Treatment System (AMTS) for On-site Destruction of PCBs Xiaoyu Liu and Zhishi Guo U.S. Environmental Protection Agency Office of Research and Development National Risk Management Research Laboratory Air Pollution Prevention and Control Division Research Triangle Park, NC 27711 and Corey A. Mocka, R. Andy Stinson, Nancy F. Roache, and Joshua A. Nardin ARCADIS, US Inc. 4915 Prospectus Dr., Suite F Durham, NC 27709 NOTICE This document has been reviewed internally and externally in accordance with the U.S. Environmental Protection Agency policy and approved for publication. Mention of trade names or commercial products does not constitute endorsement or recommendation for use. Executive Summary E.1 Background Polychlorinated biphenyls (PCBs) were once used as a plasticizer in certain building materials such as caulking, sealants, and paints from the 1950s through the late 1970s. Because PCBs have a variety of adverse health effects in animals and human, federal regulations have specific requirements for use and disposal of PCB-containing materials (U.S. EPA, 2005; 2009). Briefly, building materials that contain 50 ppm or more PCBs are not authorized for use and must be disposed of as PCB bulk product waste according the Code of Federal Regulations 40 CFR §761.3 and §761.62. If PCBs have contaminated either the surrounding building materials or adjacent soil, these materials are considered PCB remediation waste, which is subject to the cleanup and disposal requirements according 40 CFR §761.61. -

Use the Right Citrus-Based Cleaning Products to Avoid Corrosion Or Rust Bob Beckley, Project Leader
United States Department of Agriculture Facilities Forest Service Technology & Development Program March 2006 0673–2319–MTDC 7300/7100/5100/2400/2300 Use the Right Citrus-Based Cleaning Products to Avoid Corrosion or Rust Bob Beckley, Project Leader itrus-based cleaning products are commonly found in metal on their chain saws. The crew stopped using citrus-based residential and commercial settings. The ingredients in products because they believed citric acid was causing the these products vary widely (figure 1). While some of damage. However, the damage probably was caused by a C water-based citrus cleaning product. What To Look for in a Citrus-Based Cleaning Product The Material Safety Data Sheets (MSDSs) for chemical products list their ingredients. The MSDS for a citrus-based cleaner should list D-Limonene among the ingredients. D- Limonene is in the terpene family, which includes citrus and pine oils. Terpenes are generally not corrosive or harmful to metals or most plastics and polymers. Terpenes won’t cause rusting, pitting, etching, or staining. Citrus-based terpenes can dissolve heavy petroleum greases and residues in about 30 Figure 1—Citrus-based cleaners are commonly used in residential and minutes when they are used at ambient temperatures. commercial settings, but users often are unaware of the difference between citrus oil-based cleaning products and water-based products. A citrus oil-based cleaning product will not cause corrosion these products can cause corrosion or rust, others do not. The or rust. Such products are made from the oil found in the difference is based on the ingredients. Hundreds of cleaning orange peel, rather than the pulp and juice of the orange. -

Part I. Isolation of (R)-(+)-Limonene from Grapefruit Or Orange Peel
EXPERIMENT 5 ISOLATION OF LIMONENE FROM NATURAL PRODUCTS: STEAM DISTILLATION AND SOXHLET EXTRACTION OF CITRUS PEELS & MINT LEAVES Part 1: Isolation of limonene from citrus peel by steam distillation Part 2: Isolation of limonene from mint leaves by soxhlet extraction Students will work in teams of two. Each team will do one steam distillation, and one soxhlet extraction. If there is an odd number of students, then the student will do either a steam distillation or extraction, depending on availability of glassware. Reading Assignment: Pavia, Sections 18.1-18.4, 12.12 Pre-lab Questions: 1) Questions #1, 2, Pavia p. 284 - 285 2) In the spearmint leaf extraction, indicate which of the following solvents may or may not be suitable as a replacement for the hexane used in the experiment: acetone, diethyl ether, ethanol. Briefly explain your choices. Part I. Isolation of (R)-(+)-Limonene from Grapefruit or Orange Peel: (R)-(+)-limonene Line and 3-D structures of (R)-(+)-limonene. The major constituent of the steam-volatile oil of grapefruit or orange peel is (R)-(+)-limonene. It can be isolated as an esential oil of about 97% purity by a steam distillation of citrus fruit peels. Attempting to distill this essential oil directly from plant CHEM M52LA/H52LA Experiment 5 Page 1 763 material is generally not feasible. Limonene is a high-boiling liquid (bp = 175.5-176 °C) which decomposes under the high heat needed to bring it to a boil. Steam distillation is a much gentler way to obtain essential oils, as it allows the distillation of co-mixture of oil and water at a boiling point less than 100 °C. -

Wöhler Synthesis of Urea
Wöhler synthesis of urea Wöhler, 1928 – – + + NH Cl + K N C O 4 NH4 N C O O H2N NH2 Friedrich Wöhler Annalen der Physik und Chemie 1828, 88, 253–256 Significance: Wöhler was the first to make an organic substance from an inorganic substance. This was the beginning of the end of the theory of vitalism: the idea that organic and inorganic materials differed essentially by the presence of the “vital force”– present only in organic material. To his mentor Berzelius:“I can make urea without thereby needing to have kidneys, or anyhow, an animal, be it human or dog" Friedrich Wöhler, 1880-1882 Polytechnic School in Berlin Support for vitalism remained until 1845, when Kolbe synthesized acetic acid Polytechnic School at Kassel from carbon disulfide University of Göttingen Fischer synthesis of glucose PhHN N O N O NHPh Br aq. HCl Δ PhNH2NH2 HO H HO H O Br "α−acrose" H OH H OH H OH H OH CH2OH CH2OH α−acrosazone α−acrosone an “osazone” (osazone test for reducing sugars) Zn/AcOH OH OH CO2H CHO O HO H HO H OH Br2 HO H HO H Na-Hg HO H HNO3 HO H H OH H OH H OH H OH then resolve H O+ H OH via strychnine H OH H OH 3 H OH CH2OH salts CH2OH CH OH CH2OH 2 D-Mannonic acid DL-Mannose DL-Mannitol DL-Fructose quinoline • Established stereochemical relationship CO2H CHO H OH H OH between mannose and glucose Na-Hg HO H HO H (part of Fischer proof) H OH + H OH H3O • work mechanisms from acrosazone on H OH H OH Emil Fischer, 1852-1919 CH2OH CH2OH University of Munich (1875-81) D-Gluconic acid D-Glucose University of Erlangen (1881-88) University of Würzburg (1888-92) University of Berlin (1892-1919) Fischer, E. -

Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives
plants Article Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives Clémentine Baccati 1, Marc Gibernau 1, Mathieu Paoli 1 , Patrick Ollitrault 2,3 ,Félix Tomi 1,* and François Luro 2 1 Laboratoire Sciences Pour l’Environnement, Equipe Chimie et Biomasse, Université de Corse—CNRS, UMR 6134 SPE, Route des Sanguinaires, 20000 Ajaccio, France; [email protected] (C.B.); [email protected] (M.G.); [email protected] (M.P.) 2 UMR AGAP Institut, Université Montpellier, CIRAD, INRAE, Institut Agro, 20230 San Giuliano, France; [email protected] (P.O.); [email protected] (F.L.) 3 CIRAD, UMR AGAP, 20230 San Giuliano, France * Correspondence: [email protected]; Tel.: +33-495-52-4122 Abstract: The Papeda Citrus subgenus includes several species belonging to two genetically distinct groups, containing mostly little-exploited wild forms of citrus. However, little is known about the potentially large and novel aromatic diversity contained in these wild citruses. In this study, we characterized and compared the essential oils obtained from peels and leaves from representatives of both Papeda groups, and three related hybrids. Using a combination of GC, GC-MS, and 13C-NMR spectrometry, we identified a total of 60 compounds in peel oils (PO), and 76 compounds in leaf oils (LO). Limonene was the major component in almost all citrus PO, except for C. micrantha and C. hystrix, where β-pinene dominated (around 35%). LO composition was more variable, with different Citation: Baccati, C.; Gibernau, M.; major compounds among almost all samples, except for two citrus pairs: C. -

Chemical Composition, Antimicrobial, Antioxidant, and Antiproliferative Properties of Grapefruit Essential Oil Prepared by Molecular Distillation
molecules Article Chemical Composition, Antimicrobial, Antioxidant, and Antiproliferative Properties of Grapefruit Essential Oil Prepared by Molecular Distillation Weihui Deng, Ke Liu, Shan Cao, Jingyu Sun, Balian Zhong and Jiong Chun * National Navel Orange Engineering Research Center, College of Life Sciences, Gannan Normal University, Ganzhou 341000, China; [email protected] (W.D.); [email protected] (K.L.); [email protected] (S.C.); [email protected] (J.S.); [email protected] (B.Z.) * Correspondence: [email protected]; Tel.: +86-797-839-3068 Academic Editor: Raffaele Capasso Received: 3 December 2019; Accepted: 3 January 2020; Published: 5 January 2020 Abstract: Grapefruit essential oil has been proven to have wide range of bioactivities. However, bioactivity of its molecular distillate has not been well studied. In this study, a light phase oil was obtained by molecular distillation from cold-pressed grapefruit essential oil and GC-MS was used to identify its chemical composition. The antimicrobial activity of the light phase oil was tested by filter paper diffusion method, and the anticancer activity was determined by the Cell Counting Kit-8 (CCK-8) assay. Twenty-four components were detected with a total relative content of 99.74%, including 97.48% of terpenes and 1.66% of oxygenated terpenes. The light phase oil had the best antimicrobial effect on Bacillus subtilis, followed by Escherichia coli, Staphylococcus aureus and Salmonellaty phimurium. DPPH and ABTS assays demonstrated that the light phase oil had good antioxidant activity. The CCK-8 assay of cell proliferation showed that the light phase oil had a good inhibitory effect on the proliferation of HepG2 liver cancer cells and HCT116 colon cancer cells. -
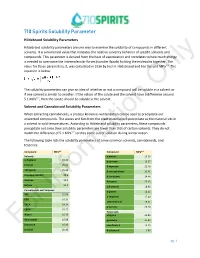
710 Spirits Solubility Parameter Hildebrand Solubility Parameters
710 Spirits Solubility Parameter Hildebrand Solubility Parameters Hildebrand solubility parameters are one way to examine the solubility of compounds in different solvents. It is a numerical value that indicates the relative solvency behavior of specific solvents and compounds. This parameter is derived from the heat of vaporization and correlates to how much energy is needed to overcome the intermolecular forces (van der Waals) holding the molecules together. The value for these parameters, δ, was calculated in 1936 by Joel H. Hildebrand and has the unit MPa1/2. The equation is below: The solubility parameters can give an idea of whether or not a compound will be soluble inOnly a solvent or if one solvent is similar to another. If the values of the solute and the solvent have a difference around 5.1 MPa1/2, then the solute should be soluble in the solvent. Solvent and Cannabinoid Solubility Parameters When extracting cannabinoids, a process known as winterization can be used to precipitate out unwanted compounds. The waxes and fats from the plant material will precipitate as the material sits in a solvent in cold temperatures. According to Hildebrand solubility parameters, these compounds precipitate out since their solubility parameters are lower than that of certain solvents. They do not match the difference of 5.1 MPa1/2 so they come out of solution during winterization. The following table lists the solubility parameters of some common solvents, cannabinoids, and terpenes: Compound MPa1/2 Compound MPa1/2 Solvents a-pinene 16.16 n-Heptane 15.40 p-cymene 16.57 Ethanol 26.20 B-myrcene 21.46 710 Spirits 25.66 B-caryophyllene 21.41 Isopropyl Alcohol 23.6 d-Limonene 24.46 Acetone 19.9 Pulegone 25.65 Butane 14.1 1,8-cineole 18.96 Cannabinoids and Terpenes a-pinene 16.15 THC Information22.09 a-terpineol 17.10 CBD 24.34 terpineol-4-ol 18.21 THCA 24.14 p-cymene 24.20 CBDA 25.77 Flavonoids Waxes 16.36 apigenin 20.80 Chlorophyll 16.98 quercetin 25.89 Limonene 16.90 cannflavin A 10.35 ForLinalool 20.46 B-sitosterol 3.86 pg.