Massive Salp Outbreaks in the Inner Sea of Chiloé Island (Southern Chile): Possible Causes and Ecological Consequences
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Región De LOS Lagos
REGIÓN DE LOCALIDADES _700 1 LOS LAGOS _conectando Chile PROYECTO { Localidades } 700_ X Proyecto _Escala Regional 108 Localidades rurales conectadas en la Región. 19.647 US$ 31,53 millones Habitantes inversión en la región. beneficiados. 42 Escuelas con servicios de Internet. LOCALIDADES _700 1 La conectividad provista por será mediante tecnología: 3G en banda 900 US$ 31,53 En el caso de la telefonía móvil será necesario uso de equipos con sello banda 900. Las escuelas contarán con servicio de Internet gratuito por dos años. “Chile está impulsando fuertemente el desarrollo de las telecomunicaciones, modernizando al país en un conjunto de ámbitos”. LOCALIDADES _700 3 Rodrigo Ramírez Pino » Subsecretario de Telecomunicaciones “Las contraprestaciones son la de las grandes urbes. Y justa, oportuna, necesaria y potente porque pone fin a la discrimina- puesta en práctica de una política ción con la que se ha tratado a pública con visión estratégica las personas que habitan esos sobre la matriz digital que Chile casi desconocidos rincones del necesita, que expresa calidad, territorio, desde el mercado de equidad y justicia digital. Cali- las telecomunicaciones”. dad, porque técnicamente se ha pensado como un acceso efectivo. Equitativa, porque iguala el acceso a la infraestructura de las telecomunicaciones en zonas aisladas del territorio respecto 4 REGIÓN DE LOS LAGOS Luisa Fuentes » Jefa de Proyectos Construcción de Red Entel “Los principales beneficios radican La experiencia ha sido motivadora en un único objetivo: hacer del país, y muy desafiante, el trabajo en uno más inclusivo en acceso a Te- equipo ha sido fundamental para lecomunicaciones, aumentando la el logro de los resultados a la eficiencia de los servicios locales, fecha. -

Acta Sesión Ordinaria Del Consejo De Monumentos Nacionales
1 MINISTERIO DE EDUCACIÓN CONSEJO DE MONUMENTOS NACIONALES ACTA SESIÓN ORDINARIA DEL CONSEJO DE MONUMENTOS NACIONALES Miércoles 10 de diciembre de 2014 2 ÍNDICE Siglas ...................................................................................................................................... 3 Apertura de la Sra. María Loreto Torres, Consejera representante del Minvu ........... 6 Informa el Sr. José de Nordenflycht Concha, Secretario Ejecutivo .............................. 7 Generales ............................................................................................................................ 10 Comisión de Arquitectura y Patrimonio Urbano ............................................................ 16 Obra menor y otros ....................................................................................................... 45 Fe de erratas ................................................................................................................... 63 Comisión de Patrimonio Histórico .................................................................................... 65 Comisión de Patrimonio Arqueológico ............................................................................ 74 Sistema de Evaluación de Impacto Ambiental ............................................................. 103 Comisión de Patrimonio Natural .................................................................................... 126 Generales (Continuación) .............................................................................................. -

Exploring the Historical Earthquakes Preceding the Giant 1960 Chile Earthquake in a Time-Dependent Seismogenic Zone
Central Washington University ScholarWorks@CWU All Faculty Scholarship for the College of the Sciences College of the Sciences 11-7-2017 Exploring the Historical Earthquakes Preceding the Giant 1960 Chile Earthquake in a Time-Dependent Seismogenic Zone Marco Cisternas Matias Carvajal Robert Wesson Lisa L. Ely Nicolás Gorigoitia Follow this and additional works at: https://digitalcommons.cwu.edu/cotsfac Part of the Geology Commons, and the Geophysics and Seismology Commons Bulletin of the Seismological Society of America, Vol. , No. , pp. –, , doi: 10.1785/0120170103 Ⓔ Exploring the Historical Earthquakes Preceding the Giant 1960 Chile Earthquake in a Time-Dependent Seismogenic Zone by M. Cisternas, M. Carvajal, R. Wesson, L. L. Ely, and N. Gorigoitia Abstract New documentary findings and available paleoseismological evidence provide both new insights into the historical seismic sequence that ended with the giant 1960 south-central Chile earthquake and relevant information about the region’s seismogenic zone. According to the few available written records, this region was previously struck by earthquakes of varying size in 1575, 1737, and 1837. We ex- panded the existing compilations of the effects of the two latter using unpublished first-hand accounts found in archives in Chile, Peru, Spain, and New England. We further investigated their sources by comparing the newly unearthed historical data and available paleoseismological evidence with the effects predicted by hypothetical dislocations. The results reveal significant differences in the along-strike and depth distribution of the ruptures in 1737, 1837, and 1960. While the 1737 rupture likely occurred in the northern half of the 1960 region, on a narrow and deep portion of the megathrust, the 1837 rupture occurred mainly in the southern half and slipped over a wide range of depth. -

Informe Etapa 3
INFORME ETAPA 3 ANTEPROYECTO ESTUDIO “ACTUALIZACIÓN PLAN REGULADOR COMUNAL DE CHONCHI Y PLANO DE DETALLE” MEMORIA EXPLICATIVA Enero 2020 ESTUDIO ACTUALIZACIÓN PLAN REGULADOR COMUNAL DE CHONCHI Y PLANO DE DETALLE TABLA DE CONTENIDOS CAPITULO I INTRODUCCIÓN ................................................................................................ 5 I.- PRESENTACIÓN ............................................................................................................. 5 I.1 OBJETIVOS DEL ESTUDIO ...................................................................................... 5 I.2 ÁREA DE ESTUDIO .................................................................................................. 6 II.- INSERCIÓN DE LA COMUNA DE CHONCHI EN LA REGIÓN ..................................... 7 II.1 Estrategia Regional de Desarrollo (ERD) ................................................................... 7 II.2 Plan Regional de Desarrollo Urbano (PRDU) Los Lagos ........................................... 8 II.3 Plan Regional de Ordenamiento Territorial (PROT) ................................................... 9 III.- INSERCIÓN DE LA COMUNA EN LA PROVINCIA DE CHILOÉ ................................. 10 IV.- ANTECEDENTES DE HISTORIA COMUNAL ............................................................. 13 IV.1 La prehistoria ........................................................................................................... 13 IV.2 La ocupación europea ............................................................................................ -

Coversheet for Thesis in Sussex Research Online
A University of Sussex DPhil thesis Available online via Sussex Research Online: http://sro.sussex.ac.uk/ This thesis is protected by copyright which belongs to the author. This thesis cannot be reproduced or quoted extensively from without first obtaining permission in writing from the Author The content must not be changed in any way or sold commercially in any format or medium without the formal permission of the Author When referring to this work, full bibliographic details including the author, title, awarding institution and date of the thesis must be given Please visit Sussex Research Online for more information and further details The politics of system innovation for emerging technologies: Understanding the uptake of off-grid renewable electricity in rural Chile Jose Opazo A thesis submitted in September 2014 in partial fulfilment of the requirements for the degree of Doctor of Philosophy SPRU - Science Policy Research Unit University of Sussex i I hereby declare that this thesis has not been, and will not be, submitted in whole or in part to another University for the award of any other degree. Signature: ……………………………………… Jose Opazo ii UNIVERSITY OF SUSSEX Jose Opazo DPhil in Science and Technology Policy Studies The politics of system innovation for emerging technologies: understanding the uptake of off-grid renewable electricity in rural Chile Summary Access to sustainable energy in the developing world has become a fundamental challenge in development and environmental policy in the 21st Century, and rural electrification in developing countries constitutes a central element of access to energy goals. However, traditional ways of providing electricity to dispersed rural populations (i.e. -

Isles of Myths&Monstersseakayak
ExChile Greatest Playground on Earth! 2009-2010 season Trip Name: Isle of Myths and Monsters Sea Kayaking Last Name: First Name: Email: Phone: # in group: Comments: Overview: Immerse yourself in the fascinating, cultural experience of Chiloe Island as we sea kayak silently through the mysterious byways of this archipelago, where Spain once maintained it's last stronghold of colonial power in South America. Here the magic of traditional culture lives on. Our sturdy kayaks emulate the spirit of the fragile dalcas; in which the natives traveled many centuries ago. Today there still vestiges of this original Araucanian way of life, in harmonious combination with the influences of German immigrants in the late 1800s and of modern-day Chile. On every lovely island we visit, you'll see moss-covered old wooden churches gracing the seaside, as well as palafitos (stilt houses) that are 20 feet and more in the air when the tide is out, but nearly awash when it's in. Savor a rich curanto and other typical cuisine to delight your palate. Wander through the local market of Castro. What more can we say? This is the opportunity of a lifetime to enjoy a fantastic adventure at the other end of the Earth National Geographic International Adventurist: Seven days to the Futaleufu - Trailer Slideshows: Slide Shows Day by Day: Day 1: ANCUD - PUÑIHUIL - CASTRO Reception at the bus terminal of Ancud, then we will drive west to visit the place of Puñihuil in the Pacific side of the main island. This is a group of islands that serve as breeding habitat for Magellanic and Humboldt Penguins as well as many other marine birds. -

Mean, Seasonal and Interannual Variability, with a Focus on 2014–2016
NASA Public Access Author manuscript Prog Oceanogr. Author manuscript; available in PMC 2020 November 16. Published in final edited form as: NASA Author ManuscriptNASA Author Manuscript NASA Author Prog Oceanogr. 2019 Manuscript NASA Author March ; 172: 159–198. doi:10.1016/j.pocean.2019.01.004. Ocean circulation along the southern Chile transition region (38° −46°S): Mean, seasonal and interannual variability, with a focus on 2014–2016 P. Ted Struba,*, Corinne Jamesa, Vivian Montecinob, José A. Rutllantc,d, José Luis Blancoe aCollege of Earth, Ocean, and Atmospheric Sciences, Oregon State University, 104 CEOAS Admin. Bldg, Corvallis, OR 97331-5503, United States bDepartamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Casilla 653, Santiago, Chile cDepartamento de Geofísica, Facultad de Ciencias Físicas y Matemáticas, Universidad de Chile, Casilla 2777, Santiago, Chile dCenter for Advanced Studies in Arid Zones (CEAZA), Coquimbo, Chile eBluewater Consulting Company, Ramalab Laboratory, O’Higgins 464, Castro, Chile Abstract Satellite and atmospheric model fields are used to describe the wind forcing, surface ocean circulation, temperature and chlorophyll-a pigment concentrations along the coast of southern Chile in the transition region between 38° and 46°S. Located inshore of the bifurcation of the eastward South Pacific Current into the equatorward Humboldt and the poleward Cape Horn Currents, the region also includes the Chiloé Inner Sea and the northern extent of the complex system of fjords, islands and canals that stretch south from near 42°S. The high resolution satellite data reveal that equatorward currents next to the coast extend as far south as 48°−51°S in spring- summer. -
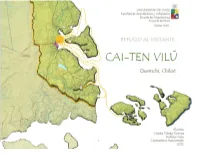
Toledo C.Pdf
1 UNIVERSIDAD DE CHILE Facultad de Arquitectura y Urbanismo Escuela de Arquitectura MEMORIA DE PROYECTO DE TÍTULO Refugio al visitante CAI -TEN VILÚ Chiloé. Arquitecto y Profesor Guía Alumna: Claudia Toledo Guevara Otoño 2012 2 3 INDICE. Asesores Profesionales 5 Motivaciones 9 ANTECEDENTES TERRITORIALES PROYECTO PRESENTACION Análisis territorial 41 Chiloé 43 Propuesta territorial 59 Extracto 11 Chiloé: tradición y cultura de un pueblo de Quemchi poseedora de un gran Tema 12 Borde mar 44 Maritorio 60 Metodología 14 Elección del lugar 46 Conectividad 6 2 Problemática 15 Quemchi 48 Propuesta territorial 64 Proyecto 16 Reseña histórica 49 Propuesta conceptual 66 Referentes 18 Riquezas de Quemchi 52 Propuesta Arquitectónica 68 Paisaje Quemchino 53 Paisaje 69 ANTECEDENTES GENERALES Quemchi y sus tradiciones 54 Físico – espacial 70 Quemchi y la madera 55 Programa social y terrestre 71 Desarrollo sustentable 21 Quemchi agrícola 56 Programa marítimo 72 Desarrollo sustentable arq. y paisaje 22 Quemchi y el mar 57 Diseño y clima 73 Turismo Nacional 24 Proyecto y sus soluciones para el clima 80 Turismo sustentable 29 Proyecto funcionamiento 82 Tipos de turismo según su aporte 30 Usuario 84 Impacto de la act. Turística 31 Normativas 86 Turismo sustentable en Chile 34 Financiamiento y gestión 87 Turismo sustentable en la Región de los lagos 35 ANTECEDENTES BIBLIOGRÁFICOS. Turismo sustentable en Chiloé 37 Turismo y comunidad 38 Bibliografía 89 Conclusión 39 4 Asesores Profesionales: Jonás Retamal Arquitecto de Chiloé Polli Vera Arquitecta master en restauración arquitectónica. Chiloé Etapa Histórica Edward Rojas Arquitecto de Chiloé Etapa Construcción Luis Goldsack Arquitecto Etapa Bioclimática Jeannette Roldan Diplomada en Arquitectura sustentable Juan Carlos Garcés Pinochet Arquitecto, Doctor en Bioclimática. -

THE PEOPLES and LANGUAGES of CHILE by DONALDD
New Mexico Anthropologist Volume 5 | Issue 3 Article 2 9-1-1941 The eoplesP and Languages of Chile Donald Brand Follow this and additional works at: https://digitalrepository.unm.edu/nm_anthropologist Recommended Citation Brand, Donald. "The eP oples and Languages of Chile." New Mexico Anthropologist 5, 3 (1941): 72-93. https://digitalrepository.unm.edu/nm_anthropologist/vol5/iss3/2 This Article is brought to you for free and open access by the Anthropology at UNM Digital Repository. It has been accepted for inclusion in New Mexico Anthropologist by an authorized editor of UNM Digital Repository. For more information, please contact [email protected]. 72 NEW MEXICO ANTHROPOLOGIST THE PEOPLES AND LANGUAGES OF CHILE By DONALDD. BRAND This article initiates a series in which the writer will attempt to summarize the scattered and commonly contradictory material on the present ethnic and linguistic constituency of a number of Latin Ameri- can countries. It represents some personal investigations in the field and an examination of much of the pertinent literature. Chile has been a sovereign state since the War of Independence 1810-26. This state was founded upon a nuclear area west of the Andean crest and essentially between 240 and 460 South Latitude. Through the War of the Pacific with Bolivia and Perui in 1879-1883 and peaceful agreements with Argentina, Chile acquired her present extention from Arica to Tierra del Fuego. These northern and south- ern acquisitions added little to her population but introduced numerous small ethnic and linguistic groups. Chile has taken national censuses in 1835, 1843, 1854, 1865, 1875, 1885, 1895, 1907, 1920, 1930, and the most recent one in November of 1940. -

Organic Matter Distribution, Composition and Its Possible Fate in the Chilean North-Patagonian Estuarine System
Science of the Total Environment 657 (2019) 1419–1431 Contents lists available at ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv Organic matter distribution, composition and its possible fate in the Chilean North-Patagonian estuarine system Humberto E. González a,b,⁎, Jorge Nimptsch a, Ricardo Giesecke a,b,NelsonSilvac a Universidad Austral de Chile, Instituto de Ciencias Marinas y Limnológicas, Valdivia, Chile b Centro FONDAP de Investigación de Ecosistemas Marinos de Altas Latitudes (IDEAL), Universidad Austral de Chile, Valdivia, Chile c Escuela de Ciencias del Mar, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile HIGHLIGHTS GRAPHICAL ABSTRACT • The distribution, composition and transport of both DOC and POC, were studied across a terrestrial-marine transition system. • A fDOM land-ocean gradient from humic-C1 terrigenous-derived to tyrosine-like C3 autochthonous fDOM was observed. • The UVC to UVA humic ratio was correlated with salinity, highlighting the high variability in fDOM chemical characteristics. • Climate warming and anthropogenic practices boost the mobilization of terrestrial carbon pools. article info abstract Article history: The distribution, composition, and transport of both dissolved and particulate organic carbon (DOC and POC) Received 25 June 2018 were studied across a terrestrial - marine transition system in the Chilean North-Patagonia (41°S). Received in revised form 26 November 2018 At the land-fjord boundary we reported: (i) high concentrations of both silicic acid (up to 100 μM) and integrated Accepted 29 November 2018 − chlorophyll a (62 mg m 2), (ii) dominance of nanophytoplankton (63%), humic-, terrigenous-derived, and Available online 04 December 2018 protein-like DOC (19 and 36%, respectively), and (iii) a shallow photic zone (12 m depth). -

Comuna Dalcahue, Atractivos Turísticos
COMUNA DALCAHUE, ATRACTIVOS TURÍSTICOS ABRIL DE 2020 INTRODUCCIÓN El turismo, es una actividad transversal al desarrollo de cualquier comuna, región o país. Es una de las áreas económicas con mayor crecimiento en los últimos años. En este sentido, Chile y en específico la comuna de Dalcahue, cuentan con un importante potencial de recursos y atractivos naturales y culturales de interés turístico, los que, junto a una oferta de servicios y demanda cada vez más creciente, constituyen una oportunidad de decisión al momento de invertir. El presente documento tiene como objetivo identificar los sectores de la comuna que destacan por su importancia turística. 1 I. ATRACTIVOS TURÍSTICOS 2 1.1 Alerzales del Fundo Potrero Anay (SN) 1.2 Astilleros de Dalcahue Éste atractivo turístico, fue declarado Santuario de la Naturaleza Es la comunidad más antigua de la comuna de Dalcahue. el 1 de septiembre de 1976. Los alerzales se encuentran Antiguamente fue un lugar de construcción de embarcaciones, ubicados en el macizo montañoso, denominado Cordillera de actividad valiosa por tratarse de estar ubicada en un Piuchén, en la zona higromórfica (Pizano y Fuenzalida, 1950), en archipiélago, que, por su geografía, las comunicaciones más la formación vegetal denominada Selva Valdiviana Andina, de relevantes se realizaron a través del mar. En los siglos XVIII y XIX vegetación mixta y correspondiente a un clima templado frío. siguió siendo importante para la carpintería, destiladeros de alcohol, actividades forestales, artesanales y campesinas. El alerce (Fitzroya cupressoides) en ocasiones, se encuentra formando una población pura o asociado con coigüe Los astilleros se ubican en la ruta Dalcahue-Castro por la costa, (Nothofagus dombeyi), tineo (Weinmannia trichosperma), mañío donde destaca la presencia de una iglesia que fue trasladada en macho (Podocarpus nubigena) y canelo (Drimys winteri). -

El Caso De Buzos Y Pescadores De Isla Guafo, Patagonia Chilena
UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS POLÍTICAS Y SOCIOLOGÍA TESIS DOCTORAL Hombres, imaginarios y paisajes, la construcción social del espacio habitado: el caso de buzos y pescadores de Isla Guafo, Patagonia chilena MEMORIA PARA OPTAR AL GRADO DE DOCTOR PRESENTADA POR Iñaki Moulian Jara Directores José Carmelo Lisón Yanko González Madrid © Iñaki Moulian Jara, 2020 UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS POLÍTICAS Y SOCIOLOGÍA TESIS DOCTORAL “Hombres, imaginarios y paisajes, la construcción social del espacio habitado: el caso de buzos y pescadores de Isla Guafo, Patagonia chilena.” Presentada por Iñaki Moulian Jara. Directores: Dr. José Carmelo Lisón. Prof. Universidad Complutense de Madrid. Dr. Yanko González. Prof. Universidad Austral de Chile. 2020 UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS POLÍTICAS Y SOCIOLOGÍA TESIS DOCTORAL “Hombres, imaginarios y paisajes, la construcción social del espacio habitado: el caso de buzos y pescadores de Isla Guafo, Patagonia chilena.” Presentada por Iñaki Moulian Jara. Directores: Dr. José Carmelo Lisón. Prof. Universidad Complutense de Madrid. Dr. Yanko González. Prof. Universidad Austral de Chile. 2020 1 3 INDICE RESUMEN…………………………………………………………………………………….6 Palabras clave……………………………………………………………………………….6 ABSTRACT……………………………………………………………………………………7 Keywords…………………………………………………………………………………...7 1) INTRODUCCIÓN…………………………………………………………………………..9 2) ANTECEDENTES………………………………………………………………………...15 3) MARCO TEÓRICO………………………………………………………………..……...17 3.1) Sobre la etnografía…………………………………………………...……………….17