The Use of Testate Amoebae in Monitoring Peatland Restoration Management: Case Studies from North West England and Ireland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Walk 3 in Between
The Salford Trail is a new, long distance walk of about 50 public transport miles/80 kilometres and entirely within the boundaries The new way to find direct bus services to where you of the City of Salford. The route is varied, going through want to go is the Route Explorer. rural areas and green spaces, with a little road walking walk 3 in between. Starting from the cityscape of Salford Quays, tfgm.com/route-explorer the Trail passes beside rivers and canals, through country Access it wherever you are. parks, fields, woods and moss lands. It uses footpaths, tracks and disused railway lines known as ‘loop lines’. Start of walk The Trail circles around to pass through Kersal, Agecroft, Bus Number 92, 93, 95 Walkden, Boothstown and Worsley before heading off to Bus stop location Littleton Road Post Office Chat Moss. The Trail returns to Salford Quays from the historic Barton swing bridge and aqueduct. During the walk Bus Number 484 Blackleach Country Park Bus stop location Agecroft Road 5 3 Clifton Country Park End of walk 4 Walkden Roe Green Bus Number 8, 22 5 miles/8 km, about 2.5 hours Kersal Bus stop location Manchester Road, St Annes’s church 2 Vale 6 Worsley 7 Eccles Chat 1 more information Moss 8 Barton For information on any changes in the route please Swing Salford 9 Bridge Quays go to visitsalford.info/thesalfordtrail kersal to clifton Little Woolden 10 For background on the local history that you will This walk follows the River Irwell upstream Moss as it meanders through woodland and Irlam come across on the trail or for information on wildlife please go to thesalfordtrail.btck.co.uk open spaces to a large country park. -

Manchester M2 6AN Boyle 7 C Brook Emetery Track Telephone 0161 836 6910 - Facsimile 0161 836 6911
Port Salford Project Building Demolitions and Tree Removal Plan Peel Investments (North) Ltd Client Salford CC LPA Date: 28.04.04 Drawing No.: 010022/SLP2 Rev C Scale: 1:10 000 @Application A3 Site Boundary KEY Trees in these areas to be retained. Scattered or occasional trees within these areas to be removed SB 32 Bdy t & Ward Co Cons SL 42 Const Bdy Boro Chat Moss CR 52 Buildings to be Demolished MP 25.25 OAD B 62 ODDINGTON ROA STANNARD R Drain 9 8 72 D 83 43 5 6 GMA PLANNING M 62 36 35 SP 28 35 27 48 3 7 2 0 19 4 0 Drain C HA Drain TLEY ROAD 3 MP 25.25 6 23 King Street, Manchester M2 6AN 12 Planning and Development Consultants Chat Moss 11 CR 32 rd Bdy Wa nst & Co Co Bdy Const e-mail [email protected] o Bor 2 53 8 1 Telephone 0161 836 6910 - Facsimile 0161 836 6911 22 Barton Moss 10 16 ROAD F ETON OXHIL BRER 9 rain 43 D L ROAD 23 Drain 2 0 St Gilbert's 33 Catholic Church MP 25 Presbytery 10 3 2 2 4 Drain Barton Moss 2 Drain Drain CR Drain 1 13 15 Co Const Bdy 6 Track Barton Moss 16 Dra Boro Const and Ward Bdy in MP 24.75 27 Eccles C of E High S Drain FLEET ROAD 6 3 ORTH 26 N SL chool D rain 0 3 Drain 39 Drai n 36 Drain BUC KT HORN D E L OA R Drain AN E D ra ILEY in H M 62 53 44 51 55 Dra 5 9 0 5 in M 62 Drain Brookhouse k Sports Centre Barton Moss Primary School rac T 0 6 63 H ILEY ROA D 6 N 5 O BU RTH D 78 rai 2 CK FLEE n T 67 4 35 H O 3 3 54 RN LA 6 T Pavilion 3 ROAD 75 N 74 E 34 27 25 18 78 6 20 7 80 88 Drain 1 1 TRIPPIER ROAD 6 56 23 58 0 30 6 3 1 n 32 55 89 9 2 7 Drai 6 1 9 6 93 64 3 2 15 95 59 ROCHFORD R 59 2 9 15 66 -

Cadishead Viaduct and the Cheshire Lines Rail Line
Proposal to re-open Cadishead Viaduct and the Cheshire Lines rail line The art of the possible! This document has been produced by Hamilton Davies Trust (HDT) to provide background and scope for the Cadishead Viaduct potential re-opening debate. HDT has been heavily involved in the regeneration of Irlam and Cadishead since 2008, working with key local, regional and national stakeholders to make Irlam and Cadishead an even better place to live, work and enjoy. As part of our regeneration agenda, the poor state and appearance of Cadishead Viaduct has been a topic of conversation for some time. Two years ago we started to investigate the possibility of improving its appearance and putting the bridge back into use. This document summarises the history of the bridge and the compelling opportunities re-opening the viaduct would bring. HDT has now adopted the project and is championing the reopening of the viaduct for multi modal use including a new section of the national cycleway, footpath and heritage railway amongst other options. Cadishead Viaduct Built in 1893, Cadishead Viaduct is a magnificent structure which dominates the landscape across the Manchester Ship Canal. Originally constructed to run the Cheshire Lines railway over the newly built canal, the viaduct helped to service the busy factories in the area, transporting raw materials and finished goods, as well as some passenger services. Due to the decline in industry, the rail line closed to passengers in 1965 and eventually to freight in 1984 when coal exports ceased. With the line no longer in use and the high cost of maintenance, the bridge was eventually closed and industrial containers positioned at either side to prevent access to the structure. -

The Development of the Railway Network in Britain 1825-19111 Leigh Shaw-Taylor and Xuesheng You 1
The development of the railway network in Britain 1825-19111 Leigh Shaw-Taylor and Xuesheng You 1. Introduction This chapter describes the development of the British railway network during the nineteenth century and indicates some of its effects. It is intended to be a general introduction to the subject and takes advantage of new GIS (Geographical Information System) maps to chart the development of the railway network over time much more accurately and completely than has hitherto been possible. The GIS dataset stems from collaboration by researchers at the University of Cambridge and a Spanish team, led by Professor Jordi Marti-Henneberg, at the University of Lleida. Our GIS dataset derives ultimately from the late Michael Cobb’s definitive work ‘The Railways of Great Britain. A Historical Atlas’. Our account of the development of the British railway system makes no pretence at originality, but the chapter does present some new findings on the economic impact of the railways that results from a project at the University of Cambridge in collaboration with Professor Dan Bogart at the University of California at Irvine.2 Data on railway developments in Scotland are included but we do not discuss these in depth as they fell outside the geographical scope of the research project that underpins this chapter. Also, we focus on the period up to 1911, when the railway network grew close to its maximal extent, because this was the end date of our research project. The organisation of the chapter is as follows. The next section describes the key characteristics of the British transport system before the coming of the railways in the nineteenth century. -

For Sale by Private Treaty Chat Moss Herb Farm
For Sale by Private Treaty Chat Moss Herb Farm, Barton Moss Road, Eccles, Manchester M30 7RL An excellently situated detached dwellinghouse on Barton Moss. Set in open countryside and fronting Barton Moss Road, the property briefly comprises a brick built former farmhouse with flexible living accommodation, a detached garage both set in mature gardens and with a field to the rear. The whole extends to 4.71 acres or thereabouts. It is convenient for Manchester City Centre and the motorway network. The property lies approximately 3 miles west of the Trafford Centre between Eccles and Irlam just north of the M62 motorway approximately 1 mile from Barton Aerodrome. t: 01772 882277 Burlington House, 10-11 Ribblesdale Place, Preston, Lancashire, PR1 3NA. e: [email protected] www.pwcsurveyors.co.uk Directions Bedroom 4 (3.60m x 4.39m) with ensuite half tiled walls; From the west at Junction 21 of the M6 motorway, travel 1 radiator. east along the A57 towards Manchester for Family Bathroom (3.76m x 2.59m) with shower; panel approximately 7 miles. About half a mile before Barton bath; WC; wash basin; 1 radiator; half tiled walls. Aerodrome, turn left into Barton Moss Road, proceed over the M62 motorway, turn right then left. Chat Moss Buildings Herb Farm is on the left approximately 250 metres to the Detached Garage (7.58m x 6.17m) brick built with electric north. up and over double doors; concrete floor. From Eccles at Peel Green junction, follow the signs to Land Irlam along the A57 in a westerly direction. Barton The land comprises a single level field classified as being Aerodrome is on the right after about a mile and Barton Grade 1. -

Winton, Greater Manchester
Winton, Greater Manchester Coordinates: 53°29′10″N 2°21′54″W / 53.486°N Road, and St. Mary Magdalene’s Parish Church on Gras- 2.365°W mere Crescent/Westbourne Road ( Grade II listed build- ing). There is a Baptist church (although its old building Winton, originally “Withington”[1] is an area north-west of Eccles in Salford, Greater Manchester, England. The in Parrin Lane is now closed). The oldest building is Mag- dalene Centre, formerly a school before Westwood Park population of the City of Salford ward at the 2011 census was 12,067.[2] was opened, dating from 1888. Historically in Lancashire, Winton is a residential area surrounded by Patricroft, Peel Green, Monton, Barton- 2.4 Transport upon-Irwell, Eccles and Worsley. 2.4.1 Buses 1 Geography and administration First Greater Manchester and Arriva North West oper- ate bus services through Winton from Eccles town cen- Winton is between Monton, Worsley, Peel Green and tre, Manchester, the Trafford Centre and Wigan. Win- Patricroft, divided by the motorway interchange of the ton is also a through-route for buses to the Trafford M602, M60 and M62. The boundaries of Winton are Centre, Wigan, Worsley and Eccles. The following Ar- the Liverpool-Manchester railway on New Lane (borders riva Routes service Winton: 61, 62 (Eccles Circulars) with Peel Green), the railway on Worsley Road (borders and the 66 Clifton-Salford Royal Hospital. The follow- with Patricroft), the Bridgewater Canal bridge at the top ing First Manchester Routes Serve Winton: 68 Trafford of Parrin Lane (borders with Monton) and the Worsley Centre-Bolton (Little Hulton/Farnworth after 18:00), 33 Road/Barton Road change (borders with Worsley). -

Clifton Moor Care Home Fascia Design by Shack Architecture
Care Home, Greater Manchester 10% Assured Returns for 10 Years Purchase from £36,000 cash input Clifton Moor Care Home Fascia design by Shack Architecture Introducing... Clifton Moor Care Home Clifton Moor Care Home offers the opportunity for Highlights individuals and businesses to purchase bedrooms within this • 35% SOLD OUT in 10 days highly sought after high quality nursing and dementia facility • 10% Assured returns for 10 years in Greater Manchester. With 70 beds, Clifton Moor will • Purchase Price £90,000 quickly become a necessity for local authorities and primary • Optional 125% (£112,500) developer buy-back after care commission teams. 10 years As an exclusive offer to Wealth Invest clients, the developer has agreed to increase the deferred funding option to 60% of Exclusive Offer for Wealth Invest Clients the purchase price allowing for individuals to purchase whole • Rooms available for only £36,000 cash input rooms from as little as £36,000 cash input. • Interest free developer deferred payments up to £54,000 2 3 Wealth Invest ROI Exclusive Limited Offer Wealth Invest are able to offer their clients a higher level of developer Calculator As an exclusive offer to Wealth Invest deferred payment for a limited time clients, each room at Clifton Moor only. Years Cash In Assured Returns Developer Deferred Balance ROI (Return on Investment) is calculated using the Care Home is available with developer (10%)* Payments This enables clients to purchase for a following formula: deferred payments for up to £54,000. £36,000 lower cash input and allows for a much (Returns from purchase + buyback - Cost of purchase) This means you can now purchase a higher overall Return On Investment. -
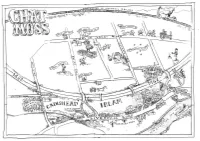
Chat-Moss-Story-Map.Pdf
Dennis Dixon Eddie Bickerton John Hannan Gwen Hall wasn’t there; rabbit pie and stews and things!’’ and stews and pie rabbit there; wasn’t or anything but we never starved because there was always rabbit rabbit always was there because starved never we but anything or Talks about working on the Chat Moss farms Talks about how his father worked with the night soil that Talks about how his great grandfather from Describes some of the difficulties the farmers But it was a good life, we enjoyed it. We weren’t massively rich rich massively weren’t We it. enjoyed we life, good a was it But as a child… fertilised and prepared Chat Moss as farmland… Ireland ended up farming on the moss... used to face… game keeping which he had to learn from scratch more or less. less. or more scratch from learn to had he which keeping game had to give up a job he liked, which was butchering and turn to to turn and butchering was which liked, he job a up give to had ‘’If we were going to school camp we used to ‘’The old chap [Eddie’s father] during the days he used ''Back in the late 1800's my great grandfather ‘’The farmers were doing quite well. This grandma and the sisters to stay in Keepers Cottage. So my dad dad my So Cottage. Keepers in stay to sisters the and grandma have to earn our own money so we used to to lean over on a steam crane and lift the night soil out of came over from Ireland to work on the was in the early 60’s. -

The Freight Solution to the Castlefield Problem
The Freight Solution to the Castlefield Problem Including The Business Case for a new route and western access to Trafford Park April 2020 (Revised August 2020) Contents Part 1. The Castlefield Problem – Freight’s Great Opportunity .................................................................................... 2 The Problem ............................................................................................................................................................... 2 A more fundamental question ................................................................................................................................... 5 Carrington Park .......................................................................................................................................................... 6 The search for a route to the south ........................................................................................................................... 7 Connecting to the West Coast Main Line .................................................................................................................. 9 Benefits of the proposed interventions ................................................................................................................... 14 Part 2. The Business Case for a Western Route to Trafford Park ................................................................................ 15 Assumptions ............................................................................................................................................................ -

How to Accommodate Forcast Growth on the Cheshire Line Corridor
How to accommodate forecast growth on the Cheshire Line Committee (CLC) corridor? Railway investment choices October 2019 02 Contents Part A: Executive Summary 03 Part B: The Long Term Planning Process and Continuous 05 Modular Strategic Planning Part C: Today’s Railway 07 Part D: Factors influencing change 13 Part E: Impact of future year growth 16 Part F: Approach to option development 19 Part G: Emerging Strategic Advice 22 Part H: Options and Advice to Funders 24 How to accommodate forecast growth on the Cheshire Line Committee (CLC) corridor? October 2019 03 Part A Executive Summary We are pleased to present an assessment of some current network assets, whilst accommodating the possible investment choices for the Cheshire Lines franchise commitments using both the CLC route and Committee (CLC) corridor between Liverpool and elsewhere. Network Rail has worked collaboratively Manchester via Warrington Central. These choices are with rail industry colleagues to consider the presented to understand which interventions may be investment choices that may be required to support required to meet future growth forecasts on the CLC this forecast growth between 2024 and 2043. corridor by 2026, 2033 and 2043. This work has been Since the development of the original report, the completed as part of the Continuous Modular economic appraisal results have been updated to Strategic Planning (CMSP) approach adopted under reflect some alternative assumptions on capital and the Long-Term Planning Process (LTPP). Industry operating costs. Whilst these revisions have improved partners have participated in the study. This the results, the updated value for money assessment collaborative approach has helped to identify some is still not sufficient to demonstrate a ‘good’ case (with possible investment choices to accommodate forecast a benefit cost ratio above 2.0). -

Script Timewalk of Chat Moss.Pdf
Timewalk of Chat Moss Close your eyes and imagine the story under your feet. The Timewalk of the 35 square kilometres of Chat Moss is incredible and starts 4 and a half billion years ago, when the earth formed. 2.7 billion years ago algae-like cells formed. They started producing oxygen, and it took 350 million years for them to make enough oxygen to turn the sky blue for the first time. More than a billion years later the first recognisable animals appeared jelly-fish and sponges in the sea. 360 million years ago the rainforest of the Carboniferous period grew which were later to form the Lancashire Coalfields. Coal formed and was buried, taking huge quantities of carbon dioxide out of the atmosphere. This cooled the planet. Coal seams were formed under Chat Moss. Continents shifted. The dinosaurs ruled the earth from 245 million years until 66 million years ago, possibly wiped out by a major asteroid that hit the earth. The Timewalk now moves forward rapidly. Our early ancestors started using stone tools 2 and a half million years ago. The earliest fossil record of modern humans is 190,000 years old. As the glaciers retreated from the Mersey Upper Terrace about 10,000 years ago, they left giant clay-lined hollows. This created nutrient-rich pools which gave way to fen carr woodland. By 3500 BCE birch trees dominated the landscape. People were beginning to settle nearby with flints found within the Woolston mosses. By 2500 BCE acid mires, within the hollows, became nutrient poor and very wet. -

Mosslands Mosslands
Lowland MOSSLANDS Lowland raised bog in the UK has fallen by 94% from 95,000 ha to 6,000 ha, and is now internationally threatened Ecology Habitat Action Plan 2008 term - raised bog. Being elevated above the Greater Manchester’s lowland mosslands, surrounding groundwater, raised bogs are (also known as lowland raised bog), began to fed purely from rainfall and this helps to form c10, 000 years ago. It dates back to the maintain nutrient poor conditions within the last ice age when peat began to be laid down bog system. The Sphagnum mosses also on marine, estuarine or fluvial deposits increase the acidity of the water. As a result, adjacent to estuaries, on river floodplains, or the characteristic vegetation found on on the site of shallow glacial lakes. These mosslands is adapted to nutrient-poor, acidic wet, waterlogged areas were originally conditions and plant species are therefore colonised by reeds and rushes. Due to the highly specialised in their requirements and waterlogged anaerobic conditions, dead plant many of these species can be found no- material could not be fully broken down and where else. began to build up on the bottom of water bodies and this led to the formation of fen Mosslands can also support characteristic peat. Bog mosses (Sphagnum mosses) assemblages of uncommon invertebrates, began to colonise. At this point, the including the large heath butterfly, once sphagnum content of the underlying peat known as the Manchester Argus, but now increased and the peat changed from fen to extinct in the County. Mossland habitat is bog peat.