Studies of the Mechanisms Involved in Host Finding And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

4Th National IPM Symposium
contents Foreword . 2 Program Schedule . 4 National Roadmap for Integrated Pest Management (IPM) . 9 Whole Systems Thinking Applied to IPM . 12 Fourth National IPM Symposium . 14 Poster Abstracts . 30 Poster Author Index . 92 1 foreword Welcome to the Fourth National Integrated Pest Management The Second National IPM Symposium followed the theme “IPM Symposium, “Building Alliances for the Future of IPM.” As IPM Programs for the 21st Century: Food Safety and Environmental adoption continues to increase, challenges facing the IPM systems’ Stewardship.” The meeting explored the future of IPM and its role approach to pest management also expand. The IPM community in reducing environmental problems; ensuring a safe, healthy, has responded to new challenges by developing appropriate plentiful food supply; and promoting a sustainable agriculture. The technologies to meet the changing needs of IPM stakeholders. meeting was organized with poster sessions and workshops covering 22 topic areas that provided numerous opportunities for Organization of the Fourth National Integrated Pest Management participants to share ideas across disciplines, agencies, and Symposium was initiated at the annual meeting of the National affiliations. More than 600 people attended the Second National IPM Committee, ESCOP/ECOP Pest Management Strategies IPM Symposium. Based on written and oral comments, the Subcommittee held in Washington, DC, in September 2001. With symposium was a very useful, stimulating, and exciting experi- the 2000 goal for IPM adoption having passed, it was agreed that ence. it was again time for the IPM community, in its broadest sense, to come together to review IPM achievements and to discuss visions The Third National IPM Symposium shared two themes, “Putting for how IPM could meet research, extension, and stakeholder Customers First” and “Assessing IPM Program Impacts.” These needs. -

The Identity of Diplospora Africana (Rubiaceae)
The identity of Diplospora africana (Rubiaceae) E. Robbrecht Nationale Plantentuin van Belgie, Domein van Bouchout, Meise, Belgium Diplospora africana Sim is shown to be a distinct species Introduction belonging to Tricalysia subg. Empogona sect. Kraussiopsis. When Sim (1907) dealt with Tricalysia in the Cape, he did It possesses the characteristics of this subgenus: Flowers not follow the delimitation of the genus proposed in with densely hairy corolla throat and appendiculate anthers, and fruits black at complete maturity. The necessary Schumann's (1891) account of the family (i.e. Diplospora and combination under the name Tricalysia and an amplified Kraussia are considered as sections of Trica/ysia) , but description of the species are provided. This rather rare distinguished between Diplospora (with tetramerous flowers), species is a Pondoland endemic separated by a wide Kraussia (pentamerous) and Tricalysia (hexamerous), stating interval from its Guineo-Congolian relatives. A key to the that this artifical distinction 'probably does not hold good species of Trica/ysia in South Africa is provided; T. africana elsewhere'. Sim recognized five species in South Africa: D . is easily distinguished from the five other southern African Trica/ysia species by its tetramerous flowers. ajricana Sim, K. lanceo/ata Sand. [ = T. lanceolata (Sand.) S. Afr. J. Bot. 1985, 51: 331-334 Burtt-Davy], K. jloribunda Harv., K. coriacea Sond. ( = T. sonderana Hiern) and T. capensis (Meisn.) Sim. While the Diplospora africana Sim is 'n maklik onderskeidbare spesie last four species are now well known elements of the South wat in Tricalysia subg. Empogona sect. Kraussiopsis African flora, Dip/ospora ajricana remained obscure and ingesluit word. -

Plant Collections from Ethiopia Desalegn Desissa & Pierre Binggeli
Miscellaneous Notes & Reports in Natural History, No 001 Ecology, Conservation and Resources Management 2003 Plant collections from Ethiopia Desalegn Desissa & Pierre Binggeli List of plants collected by Desalegn Desissa and Pierre Binggeli as part of the biodiversity assessment of church and monastery vegetation in Ethiopia in 2001-2002. The information presented is a slightly edited version of what appears on the herbarium labels (an asci-delimited version of the information is available from [email protected]). Sheets are held at the Addis Ababa and Geneva herberia. Abutilon longicuspe Hochst. ex A. Rich Malvaceae Acacia etbaica Schweinf. Fabaceae Desalgen Desissa & Pierre Binggeli DD416 Desalegn Desissa & Pierre Binggeli DD432 Date: 02-01-2002 Date: 25-01-2002 Location: Ethiopia, Shewa, Zena Markos Location: Ethiopia, Tigray, Mekele Map: 0939A1 Grid reference: EA091905 Map: 1339C2 Grid reference: Lat. 09º52’ N Long. 39º04’ E Alt. 2560 m Lat. 13º29' N Long. 39º29' E Alt. 2150 m Site: Debir and Dey Promontary is situated 8 km to the West of Site: Debre Genet Medihane Alem is situated at the edge of Mekele Inewari Town. The Zena Markos Monastery is located just below Town at the base of a small escarpment. The site is dissected by a the ridge and overlooks the Derek Wenz Canyon River by 1200 m. stream that was dry at the time of the visit. For site details go to: The woodland is right below the cliff on a scree slope. Growing on a http://members.lycos.co.uk/ethiopianplants/sacredgrove/woodland.html large rock. For site details go to: Vegetation: Secondary scrubby vegetation dominated by Hibiscus, http://members.lycos.co.uk/ethiopianplants/sacredgrove/woodland.html Opuntia, Justicia, Rumex, Euphorbia. -

4 Reproductive Biology of Cerambycids
4 Reproductive Biology of Cerambycids Lawrence M. Hanks University of Illinois at Urbana-Champaign Urbana, Illinois Qiao Wang Massey University Palmerston North, New Zealand CONTENTS 4.1 Introduction .................................................................................................................................. 133 4.2 Phenology of Adults ..................................................................................................................... 134 4.3 Diet of Adults ............................................................................................................................... 138 4.4 Location of Host Plants and Mates .............................................................................................. 138 4.5 Recognition of Mates ................................................................................................................... 140 4.6 Copulation .................................................................................................................................... 141 4.7 Larval Host Plants, Oviposition Behavior, and Larval Development .......................................... 142 4.8 Mating Strategy ............................................................................................................................ 144 4.9 Conclusion .................................................................................................................................... 148 Acknowledgments ................................................................................................................................. -

Alien Invasive Species and International Trade
Forest Research Institute Alien Invasive Species and International Trade Edited by Hugh Evans and Tomasz Oszako Warsaw 2007 Reviewers: Steve Woodward (University of Aberdeen, School of Biological Sciences, Scotland, UK) François Lefort (University of Applied Science in Lullier, Switzerland) © Copyright by Forest Research Institute, Warsaw 2007 ISBN 978-83-87647-64-3 Description of photographs on the covers: Alder decline in Poland – T. Oszako, Forest Research Institute, Poland ALB Brighton – Forest Research, UK; Anoplophora exit hole (example of wood packaging pathway) – R. Burgess, Forestry Commission, UK Cameraria adult Brussels – P. Roose, Belgium; Cameraria damage medium view – Forest Research, UK; other photographs description inside articles – see Belbahri et al. Language Editor: James Richards Layout: Gra¿yna Szujecka Print: Sowa–Print on Demand www.sowadruk.pl, phone: +48 022 431 81 40 Instytut Badawczy Leœnictwa 05-090 Raszyn, ul. Braci Leœnej 3, phone [+48 22] 715 06 16 e-mail: [email protected] CONTENTS Introduction .......................................6 Part I – EXTENDED ABSTRACTS Thomas Jung, Marla Downing, Markus Blaschke, Thomas Vernon Phytophthora root and collar rot of alders caused by the invasive Phytophthora alni: actual distribution, pathways, and modeled potential distribution in Bavaria ......................10 Tomasz Oszako, Leszek B. Orlikowski, Aleksandra Trzewik, Teresa Orlikowska Studies on the occurrence of Phytophthora ramorum in nurseries, forest stands and garden centers ..........................19 Lassaad Belbahri, Eduardo Moralejo, Gautier Calmin, François Lefort, Jose A. Garcia, Enrique Descals Reports of Phytophthora hedraiandra on Viburnum tinus and Rhododendron catawbiense in Spain ..................26 Leszek B. Orlikowski, Tomasz Oszako The influence of nursery-cultivated plants, as well as cereals, legumes and crucifers, on selected species of Phytophthopra ............30 Lassaad Belbahri, Gautier Calmin, Tomasz Oszako, Eduardo Moralejo, Jose A. -

Pests and Diseases of Coffee in East Africa
Pests and Diseases of Coffee in Eastern Africa: A Technical and Advisory Manual compiled & edited by Mike A. Rutherford CABI UK Centre (Ascot) and Noah Phiri CABI Africa Regional Centre (Nairobi) Acknowledgements The authors would like to thank all those who contributed towards the preparation of this manual, in terms of provision of information, photographic material and advice. Gratitude is also extended to the United Kingdom Department for International Development (DFID) for providing financial report through the Renewable Natural Resources Research Strategy Crop Protection Programme (RNRRS CPP). Copyright statement © Copyright CAB International (2006) Extracts of this publication may be freely reproduced and distributed on a non-commercial basis for teaching and training purposes only, providing that the source is clearly acknowledged as: CAB International (2006) Pests and diseases of coffee in eastern Africa: a technical and advisory manual. CAB International, Wallingford, UK Compiled & edited by Mike Rutherford and Noah Phiri The copyright works may not be used for any other purpose without the express written consent of CAB International (trading as CABI), and such notice shall be placed on all copies distributed by whatever means. This publication is an output from the Crop Protection Programme of the UK Department for International Development (DFID), for the benefit of developing countries. The views expressed are not necessarily those of DFID. 2 CONTENTS Page no. Part 1 Introduction 4 Part 2 Coffee Pests 7 Coffee Berry Borer -

Transcriptome and Gene Expression Analysis of Rhynchophorus Ferrugineus (Coleoptera: Curculionidae) During Developmental Stages
Transcriptome and gene expression analysis of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) during developmental stages Hongjun Yang1,2, Danping Xu1, Zhihang Zhuo1,2,3, Jiameng Hu2 and Baoqian Lu4 1 College of Life Science, China West Normal University, Nanchong, Sichuan, China 2 Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Key Laboratory of Germplasm Resources Biology of Tropical Special Ornamental Plants of Hainan Province, College of Forestry, Hainan University, Haikou, Hainan, China 3 Key Laboratory of Integrated Pest Management on Crops in South China, Ministry of Agriculture, South China Agricultural University, Guangzhou, Guangdong, China 4 Key Laboratory of Integrated Pest Management on Tropical Crops, Ministry of Agriculture China, Environ- ment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China ABSTRACT Background. Red palm weevil, Rhynchophorus ferrugineus Olivier, is one of the most destructive pests harming palm trees. However, genomic resources for R. ferrugineus are still lacking, limiting the ability to discover molecular and genetic means of pest control. Methods. In this study, PacBio Iso-Seq and Illumina RNA-seq were used to generate transcriptome from three developmental stages of R. ferrugineus (pupa, 7th-instar larva, adult) to increase the understanding of the life cycle and molecular characteristics of the pest. Results. Sequencing generated 625,983,256 clean reads, from which 63,801 full-length transcripts were assembled with N50 of 3,547 bp. Expression analyses revealed 8,583 differentially expressed genes (DEGs). Moreover, gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed that these Submitted 5 March 2020 Accepted 29 September 2020 DEGs were mainly related to the peroxisome pathway which associated with metabolic Published 2 November 2020 pathways, material transportation and organ tissue formation. -

Proceedings, 23Rd U.S. Department of Agriculture Interagency Research
United States Department of Proceedings Agriculture 23rd U.S. Department of Agriculture Forest Service Northern Interagency Research Forum on Research Station Invasive Species 2012 General Technical Report NRS-P-114 The findings and conclusions of each article in this publication are those of the individual author(s) and do not necessarily represent the views of the U.S. Department of Agriculture or the Forest Service. All articles were received in digital format and were edited for uniform type and style. Each author is responsible for the accuracy and content of his or her paper. The use of trade, firm, or corporation names in this publication is for the information and convenience of the reader. Such use does not constitute an official endorsement or approval by the U.S. Department of Agriculture or the Forest Service of any product or service to the exclusion of others that may be suitable. This publication reports research involving pesticides. It does not contain recommendations for their use, nor does it imply that the uses discussed here have been registered. All uses of pesticides must be registered by appropriate State and/or Federal, agencies before they can be recommended. CAUTION: Pesticides can be injurious to humans, domestic animals, desirable plants, and fi sh or other wildlife—if they are not handled or applied properly. Use all pesticides selectively and carefully. Follow recommended practices for the disposal of surplus pesticides and pesticide containers. Cover graphic by Vincent D’Amico, U.S. Forest Service, Northern Research Station. Manuscript received for publication August 2012 Published by: For additional copies: U.S. -

SCREENING for ANTIBACTERIAL and ANTIFUNGAL COMPOUNDS in Bersama Abyssinica FRESEN
SCREENING FOR ANTIBACTERIAL AND ANTIFUNGAL COMPOUNDS IN Bersama abyssinica FRESEN ONG’ERA TABITHA NYANCHOKA (B.ED, SC.) (I56/CE/11221/2007) A Research Thesis Submitted in Partial Fulfillment of the Requirements for the Award of the Degree of Master of Science in the School of Pure and Applied Science, Kenyatta University MAY 2016 ii Declarations I declare that this thesis is my original work and has not been previously presented for a degree in Kenyatta University or in any other University. Ong’era Tabitha Nyanchoka Reg. No. I56/CE/11221/07 Sign: _________________________Date _____________________________ Department of Chemistry We confirm that the work reported in this thesis was carried out by the candidate under our supervision. Prof. Alex K. Machocho Department of Chemistry Kenyatta University Sign: _______________________ Date_____________________________ Prof. Nicholas K. Gikonyo Department of Pharmacy, Complementary and Alternative Medicine Kenyatta University Sign: ____________________________Date__________________________ iii Dedication To my daughter Flavian Bonareri, sons Adrian Nyaenya and Fabian Ong’era and my beloved husband Evans Nyaenya. iv Acknowledgements I would like to display my sincere gratitude to the following persons for their support, assistance and encouragement throughout the course of my research and compilation of this thesis. My supervisors: Prof. Alex Machocho of the Department of Chemistry and Prof. Nicholas Gikonyo of the Department of Pharmacy, Complementary and Alternative medicine for their guidance, motivation, support and time invested in me. Also, Dr. Omari Amuka of Maseno University for assisting me in the plant collection exercise. Dr. Margret Ng’ang’a of the Department of Chemistry for her encouragement and support throughout my research in the laboratory. -

EPPO Reporting Service
ORGANISATION EUROPEENNE EUROPEAN AND ET MEDITERRANEENNE MEDITERRANEAN POUR LA PROTECTION DES PLANTES PLANT PROTECTION ORGANIZATION OEPP Service d’Information NO. 8 PARIS, 2017-08 Général 2017/145 Nouvelles données sur les organismes de quarantaine et les organismes nuisibles de la Liste d’Alerte de l’OEPP 2017/146 Liste de quarantaine de l'Union Économique Eurasiatique (EAEU) 2017/147 Kits de communication de l’OEPP : nouveaux modèles d’affiches et de brochures sur les organismes nuisibles Ravageurs 2017/148 Rhynchophorus ferrugineus n’est pas présent en Australie 2017/149 Platynota stultana (Lepidoptera : Tortricidae) : à nouveau ajouté sur la Liste d’Alerte de l’OEPP Maladies 2017/150 Premier signalement de Puccinia hemerocallidis au Portugal 2017/151 Premier signalement de Pantoea stewartii en Malaisie 2017/152 La maladie de la léprose des agrumes est associée à plusieurs virus 2017/153 Brevipalpus phoenicis, vecteur de la léprose des agrumes, est un complexe d'espèces Plantes envahissantes 2017/154 Potentiel suppressif de certaines graminées sur la croissance et le développement d’Ambrosia artemisiifolia 2017/155 Bidens subalternans dans la région OEPP : addition à la Liste d’Alerte de l’OEPP 2017/156 Les contraintes abiotiques et la résistance biotique contrôlent le succès de l’établissement d’Humulus scandens 21 Bld Richard Lenoir Tel: 33 1 45 20 77 94 E-mail: [email protected] 75011 Paris Fax: 33 1 70 76 65 47 Web: www.eppo.int OEPP Service d’Information 2017 no. 8 – Général 2017/145 Nouvelles données sur les organismes de quarantaine et les organismes nuisibles de la Liste d’Alerte de l’OEPP En parcourant la littérature, le Secrétariat de l’OEPP a extrait les nouvelles informations suivantes sur des organismes de quarantaine et des organismes nuisibles de la Liste d’Alerte de l’OEPP (ou précédemment listés). -

Program Book
NORTH CENTRAL BRANCH Entomological Society of America 59th Annual Meeting March 28-31, 2004 President Rob Wiedenmann The Fairmont Kansas City At the Plaza 401 Ward Parkway Kansas City, MO 64112 Contents Meeting Logistics ................................................................ 2 2003-2004 Officers and Committees, ESA-NCB .............. 4 2004 North Central Branch Award Recipients ................ 8 Program ............................................................................. 13 Sunday, March 28, 2004 Afternoon ...............................................................13 Evening ..................................................................13 Monday, March 29, 2004 Morning..................................................................14 Afternoon ...............................................................23 Evening ..................................................................42 Tuesday, March 30, 2004 Morning..................................................................43 Afternoon ...............................................................63 Evening ..................................................................67 Wednesday, March 31, 2004 Morning..................................................................68 Afternoon ...............................................................72 Author Index ..............................................................73 Taxonomic Index........................................................84 Key Word Index.........................................................88 -
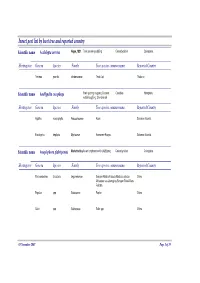
Insect Pest List by Host Tree and Reported Country
Insect pest list by host tree and reported country Scientific name Acalolepta cervina Hope, 1831 Teak canker grub|Eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Tectona grandis Verbenaceae Teak-Jati Thailand Scientific name Amblypelta cocophaga Fruit spotting bug|eng Coconut Coreidae Hemiptera nutfall bug|Eng, Chinche del Hosting tree Genera Species Family Tree species common name Reported Country Agathis macrophylla Araucariaceae Kauri Solomon Islands Eucalyptus deglupta Myrtaceae Kamarere-Bagras Solomon Islands Scientific name Anoplophora glabripennis Motschulsky Asian longhorn beetle (ALB)|eng Cerambycidae Coleoptera Hosting tree Genera Species Family Tree species common name Reported Country Paraserianthes falcataria Leguminosae Sengon-Albizia-Falcata-Molucca albizia- China Moluccac sau-Jeungjing-Sengon-Batai-Mara- Falcata Populus spp. Salicaceae Poplar China Salix spp. Salicaceae Salix spp. China 05 November 2007 Page 1 of 35 Scientific name Aonidiella orientalis Newstead, Oriental scale|eng Diaspididae Homoptera 1894 Hosting tree Genera Species Family Tree species common name Reported Country Lovoa swynnertonii Meliaceae East African walnut Cameroon Azadirachta indica Meliaceae Melia indica-Neem Nigeria Scientific name Apethymus abdominalis Lepeletier, Tenthredinidae Hymenoptera 1823 Hosting tree Genera Species Family Tree species common name Reported Country Other Coniferous Other Coniferous Romania Scientific name Apriona germari Hope 1831 Long-horned beetle|eng Cerambycidae