16 October 2013 Kevin Elliot Forest Supervisor Sequoia National Forest
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Post-Fire Treatment Effectiveness for Hillslope Stabilization
United States Department of Agriculture Post-Fire Treatment Forest Service Rocky Mountain Effectiveness for Research Station General Technical Hillslope Stabilization Report RMRS-GTR-240 August 2010 Peter R. Robichaud, Louise E. Ashmun, and Bruce D. Sims A SUMMARY OF KNOWLEDGE FROM THE Robichaud, Peter R.; Ashmun, Louise E.; Sims, Bruce D. 2010. Post-fire treatment effectiveness for hill- slope stabilization. Gen. Tech. Rep. RMRS-GTR-240. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 62 p. Abstract This synthesis of post-fire treatment effectiveness reviews the past decade of research, monitoring, and product development related to post-fire hillslope emergency stabilization treatments, including erosion barri- ers, mulching, chemical soil treatments, and combinations of these treatments. In the past ten years, erosion barrier treatments (contour-felled logs and straw wattles) have declined in use and are now rarely applied as a post-fire hillslope treatment. In contrast, dry mulch treatments (agricultural straw, wood strands, wood shreds, etc.) have quickly gained acceptance as effective, though somewhat expensive, post-fire hillslope stabilization treatments and are frequently recommended when values-at-risk warrant protection. This change has been motivated by research that shows the proportion of exposed mineral soil (or conversely, the propor- tion of ground cover) to be the primary treatment factor controlling post-fire hillslope erosion. Erosion barrier treatments provide little ground cover and have been shown to be less effective than mulch, especially during short-duration, high intensity rainfall events. In addition, innovative options for producing and applying mulch materials have adapted these materials for use on large burned areas that are inaccessible by road. -

Digitized by South Carolina State Library VOLUNTEER STRATEGIC ASSISTANCE and FIRE EQUIPMENT PILOT PROGRAM
Digitized by South Carolina State Library VOLUNTEER STRATEGIC ASSISTANCE AND FIRE EQUIPMENT PILOT PROGRAM SOUTH CAROLINA DEPARTMENT OF LABOR, LICENSING AND REGULATION DIVISION OF FIRE AND LIFE SAFETY S. C. STATE LIBRARY AUG 13 2008 STATE DOCUMENTS Certified Public Manager Project Submitted by: Sondra Senn, Manager Public Fire Education and Data Management Submitted: February 6,2008 Digitized by South Carolina State Library VOLUNTEER STRATEGIC ASSISTANCE AND FIRE EQUIPMENT PILOT PROGRAM (V-SAFE) PROBLEM STATEMENT The 2007 Session of the South Carolina General Assembly enacted the Volunteer Strategic Assistance and Fire Equipment Pilot Program Law (H3045), providing $3 million in grants to eligible volunteer and combination fire departments for the purpose of protecting local communities and regional response areas from incidents of fire, hazardous materials, terrorism, and to provide for the safety of volunteer firefighters (See Appendix A). The South Carolina Department of Labor, Licensing and Regulation, State Fire Marshal's Office, was charged with administering the grant process to include: I) developing the grant application package; 2) establishing the criteria for the evaluation of grants as outlined in the law; 3) marketing the information to fire departments; 4) developing a tracking mechanism for expenditures awarded; and 5) reporting this information back to the General Assembly at the close ofthe funding period. As identified in the legislation, funding received by fire departments must be used to purchase equipment in the following categories: fire suppression equipment; self-contained breathing apparatus; portable air filling stations; hazardous materials spill leak detection, repair and recovery equipment; protective clothing and equipment; incident command vehicles; special operations vehicles; training; rescue equipment; medical equipment, decontamination equipment; and safety equipment. -

What Information Do People Use, Trust, and Find Useful During A
Nat Hazards (2015) 76:615–634 DOI 10.1007/s11069-014-1512-x ORIGINAL PAPER What information do people use, trust, and find useful during a disaster? Evidence from five large wildfires Toddi A. Steelman • Sarah M. McCaffrey • Anne-Lise Knox Velez • Jason Alexander Briefel Received: 9 April 2014 / Accepted: 4 November 2014 / Published online: 16 November 2014 Ó Springer Science+Business Media Dordrecht 2014 Abstract The communication system through which information flows during a disaster can be conceived of as a set of relationships among sources and recipients who are concerned about key information characteristics. The recipient perspective is often neglected within this system. In this article, we explore recipient perspectives related to what information was used, useful, and trustworthy in a wildfire context. Using a survey (n = 873) on five large wildfires in 2009 and 2010, we found significant gaps between the sources that were used by the most respondents and those that that they rated as useful or trustworthy. The sources that were used most before the fires were highly correlated with the sources that were used most during the fire. Keywords Public information Á Communication Á Used Á Useful Á Trustworthy 1 Introduction Information is an essential resource during a disaster. Without information, responders cannot effectively manage a disaster, and those affected by the disaster cannot best adapt to T. A. Steelman (&) School of Environment and Sustainability, University of Saskatchewan, 329 Kirk Hall, 117 Science Place, Saskatoon, SK S7H 2J5, Canada e-mail: [email protected] S. M. McCaffrey Northern Research Station USDA Forest Service, 1033 University Place, #360, Evanston, IL 60626, USA e-mail: [email protected] A.-L. -

Fires on Flathead Forest Grow Slightly, Expected to Burn for Weeks
Houston East (CAMS 1) monitoring site AUGUST 26, 2011 EXCEPTIONAL EVENTS DEMONSTRATION PACKAGE For the Houston-Galveston-Brazoria One-hour Ozone Nonattainment Area TEXAS COMMISSION ON ENVIRONMENTAL QUALITY P.O. BOX 13087 AUSTIN, TEXAS 78711-3087 September 30, 2014 TABLE OF CONTENTS Table of Contents ................................................................................................................. i List of Acronyms ................................................................................................................ iii List of Tables ........................................................................................................................v List of Figures .................................................................................................................... vi Executive Summary ......................................................................................................... viii Chapter 1: Introduction .................................................................................................... 1-1 1.1 A Conceptual Model of Ozone for the HGB Area ................................................. 1-1 1.2 Trends in HGB Area Emissions and Ozone Levels ............................................. 1-2 1.3 Ozone Reduction Progress at the Houston East (CAMS 1) Monitoring Site ...... 1-4 1.4 The Exceptional Event of August 26, 2011 ......................................................... 1-5 Chapter 2: Exceptional Event Rule Requirements for States ......................................... -

Facing Fire Tour Checklist
EXHIBITION CHECKLIST Curated by Douglas McCulloh Organized by UCR ARTS: California Museum of Photography Noah Berger Kevin Cooley Josh Edelson Samantha Fields Jeff Frost Luther Gerlach Christian Houge Richard Hutter Anna Mayer Stuart Palley Norma I. Quintana Justin Sullivan FACING FIRE—CHECKLIST 1 Noah Berger Carr Fire, 2018 (printed 2020) Dye sublimation print on aluminum 20 x 30 inches Courtesy of the artist and Associated Press Noah Berger Delta Fire, 2018 (printed 2020) Dye sublimation print on aluminum 20 x 30 inches Courtesy of the artist and Associated Press Noah Berger Delta Fire, 2018 (printed 2020) Dye sublimation print on aluminum 24 x 36 inches Courtesy of the artist and Associated Press Noah Berger Ferguson Fire, 2018 (printed 2020) Dye sublimation print on aluminum 24 x 36 inches Courtesy of the artist and Associated Press FACING FIRE—CHECKLIST 2 Noah Berger Ferguson Fire, 2018 (printed 2020) Dye sublimation print on aluminum 24 x 36 inches Courtesy of the artist and Associated Press Noah Berger Holiday Fire, 2018 (printed 2020) Dye sublimation print on aluminum 24 x 36 inches Courtesy of the artist and Associated Press Noah Berger Justin Sullivan shooting low, Camp Fire, 2018 (printed 2020) Dye sublimation print on aluminum 24 x 36 inches Courtesy of the artist and Associated Press Noah Berger Kincade Fire, 2019 (printed 2020) Dye sublimation print on aluminum 20 x 30 inches Courtesy of the artist and Associated Press FACING FIRE—CHECKLIST 3 Noah Berger Kincade Fire, 2019 (printed 2020) Dye sublimation print on aluminum -

Collective Intelligence in Emergency Management 1
Collective Intelligence in Emergency Management 1 Running head: COLLECTIVE INTELLIGENCE IN EMERGENCY MANAGEMENT: SOCIAL MEDIA’S ROLE IN THE EMERGENCY OPERATIONS CENTER Collective Intelligence in Emergency Management: Social Media's Emerging Role in the Emergency Operations Center Eric D. Nickel Novato Fire District Novato, California Collective Intelligence in Emergency Management 2 CERTIFICATION STATEMENT I hereby certify that this paper constitutes my own product, that where the language of others is set forth, quotation marks so indicate, and that appropriate credit is given where I have used the language, ideas, expressions, or writings of another. Signed: __________________________________ Collective Intelligence in Emergency Management 3 ABSTRACT The problem was that the Novato Fire District did not utilize social media technology to gather or share intelligence during Emergency Operations Center activations. The purpose of this applied research project was to recommend a social media usage program for the Novato Fire District’s Emergency Operations Center. Descriptive methodology, literature review, two personal communications and a statistical sampling of fire agencies utilizing facebook supported the research questions. The research questions included what were collective intelligence and social media; how was social media used by individuals and organizations during events and disasters; how many fire agencies maintained a facebook page and used them to distribute emergency information; and which emergency management social media programs should be recommended for the Novato Fire District’s Emergency Operations Center. The procedures included two data collection experiments, one a statistical sampling of United States fire agencies using facebook, to support the literature review and research questions. This research is one of the first Executive Fire Officer Applied Research Projects that addressed this emerging subject. -

99921 FSPLT3 2440025.Pdf
Fire and Fuels Specialist Report-Shirley Fire Salvage 2015 Cover photo: Burned area of Shirley Fire taken near the point of origin, showing typical conditions in project area. Compare with the photo series in Appendix A which was used in analyzing the Shirley Fire project area Contents FIRE AND FUELS SPECIALIST REPORT .............................................................................................................. 1 Summary ........................................................................................................................................................... 1 Background and Affected Environment ............................................................................................................ 1 Fire Environment .......................................................................................................................................... 1 Wildland Urban Intermix .......................................................................................................................... 1 Fire Suppression ........................................................................................................................................ 1 Fire Severity and Fuel Bed Change ........................................................................................................... 2 Fire Weather, Drought, and Topography .................................................................................................. 3 Fire History ............................................................................................................................................... -

UNIVERSITY of CALIFORNIA Los Angeles Post-Fire
UNIVERSITY OF CALIFORNIA Los Angeles Post-fire hydrologic behavior and recovery: Advancing spatial and temporal prediction with an emphasis on remote sensing A dissertation submitted in partial satisfaction of the requirements for the degree Doctor of Philosophy in Civil Engineering by Alicia Michiko Kinoshita 2012 ABSTRACT OF THE DISSERTATION Post-fire hydrologic behavior and recovery: Advancing spatial and temporal prediction with an emphasis on remote sensing by Alicia Michiko Kinoshita Doctor of Philosophy in Civil Engineering University of California, Los Angeles, 2012 Professor Terri S. Hogue, Chair This work has investigated the policy of wildfires, modeling techniques for post-fire assessment, and the influence of controlling variables on post-fire recovery. Post-fire mitigation and management require reliable predictions of immediate hydrologic consequences and long-term recovery to pre-fire conditions. This research shows that models used by agencies are not adaptable to all geographical and climatological conditions. Results show inconsistencies between model predictions for peak discharge events across the sites and less confidence associated with larger return periods (25- and 50-year peak flow events). Remote sensing techniques improve spatial and temporal resolution of data streams for model parameters and post-fire recovery predictions. This research shows that recovery is dependent on many ii variables, including burn severity, slope aspect, and vegetation biomass. The lack of vegetation recovery across watersheds results in significant changes in annual and seasonal discharge throughout the study period. Understanding these key controlling variables will improve post-fire hydrological predictions. Previously established remote sensing algorithms can be applied and adapted to burned areas to improve hydrologic and recovery predictions. -

Sequoia National Forest Assessment Begins with This INTRODUCTION to Provide Background on the Process and to Describe the Assessment Area
CONTENTS INTRODUCTION ....................................................................................................................................................... 6 Purpose of the Assessment .................................................................................................................................. 6 Structure of the Assessment ................................................................................................................................ 6 Living Assessment ................................................................................................................................................... 7 Maps of the Assessment Area ............................................................................................................................. 8 Assessment Area, History and Distinctive Features ...................................................................................... 10 RESOURCES MANAGED AND EXISTING PLAN OBJECTIVES .................................................................... 12 BEST AVAILABLE SCIENTIFIC INFORMATION .............................................................................................. 16 FINDINGS ................................................................................................................................................................. 18 Chapter 1: Ecological Integrity .......................................................................................................................... 18 Important Information -
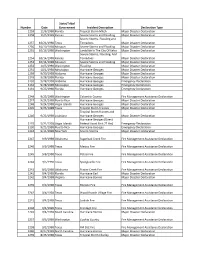
Identification of Disaster Code Declaration
State/Tribal Number Date Government Incident Description Declaration Type 1259 11/6/1998 Florida Tropical Storm Mitch Major Disaster Declaration 1258 11/5/1998 Kansas Severe Storms and Flooding Major Disaster Declaration Severe Storms, Flooding and 1257 10/21/1998 Texas Tornadoes Major Disaster Declaration 1256 10/19/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1255 10/16/1998 Washington Landslide In The City Of Kelso Major Disaster Declaration Severe Storms, Flooding, And 1254 10/14/1998 Kansas Tornadoes Major Disaster Declaration 1253 10/14/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1252 10/5/1998 Washington Flooding Major Disaster Declaration 1251 10/1/1998 Mississippi Hurricane Georges Major Disaster Declaration 1250 9/30/1998 Alabama Hurricane Georges Major Disaster Declaration 1249 9/28/1998 Florida Hurricane Georges Major Disaster Declaration 3133 9/28/1998 Alabama Hurricane Georges Emergency Declaration 3132 9/28/1998 Mississippi Hurricane Georges Emergency Declaration 3131 9/25/1998 Florida Hurricane Georges Emergency Declaration 2248 9/25/1998 Washington Columbia County Fire Management Assistance Declaration 1247 9/24/1998 Puerto Rico Hurricane Georges Major Disaster Declaration 1248 9/24/1998 Virgin Islands Hurricane Georges Major Disaster Declaration 1245 9/23/1998 Texas Tropical Storm Frances Major Disaster Declaration Tropical Storm Frances and 1246 9/23/1998 Louisiana Hurricane Georges Major Disaster Declaration Hurricane Georges (Direct 3129 9/21/1998 Virgin Islands Federal -

015-2018 SHMP FINAL Appendices
APPENDICES – 2018 STATE HAZARD MITIGATION TEAM ROSTER OF AGENCIES AND STAKEHOLDER ORGANIZATIONS AECOM National Governments Alameda County Office of Emergency Services Alpine County Operational Area Inland Region IV Air Resources Control Board (ARB) Amador County Operational Area Inland Region IV American Planning Association California Chapter American Red Cross (Sacramento Chapter) Association of Bay Area Governments Association of Contingency Planners Association of Environmental Professionals Burbank Fire Corps Business and Industry Council for Emergency Planning & Preparedness (BICEPP) Business, Consumer Services, and Housing Agency Business Executives for National Security (BENS) Business Recovery Managers Association Butte County Operational Area Inland Region III California Adaptation Forum California Association of Councils of Governments Cahuilla Band of Indians California Board of Forestry and Fire Protection Calaveras Council of Governments California Coastal Commission California Conservation Corps California Community Colleges California Department of Community Services and Development California Department of Conservation California Department of Corrections and Rehabilitation California Department of Education California Department of Food and Agriculture California Department of Forestry and Fire Protection (CALFIRE) California Department of General Services California Department of Housing and Community Development California Department of Insurance California Department of Public Health California Department of Public -

René Voss – Attorney at Law 15 Alderney Road San Anselmo, CA 94960 Tel: 415-446-9027 [email protected] ______
René Voss – Attorney at Law 15 Alderney Road San Anselmo, CA 94960 Tel: 415-446-9027 [email protected] _____________________________________________________________________________________________ Sent to: October 16, 2015 [email protected] Alfred W. Watson, District Ranger cc: Ara Marderosian Steve Anderson, Project Planner Dr. Michael Connor Kern River Ranger District Alison Sheehey PO Box 9 Joe Fontaine Kernville, CA 93238 Kevin Elliott Subject: Summit Project EIS Scoping Comments for Sequoia ForestKeeper, the Kern- Kaweah Chapter of the Sierra Club, and Western Watersheds Project Sequoia ForestKeeper, the Kern-Kaweah Chapter of the Sierra Club, and Western Watersheds Project thank you for the opportunity to comment. Background The Forest Service is planning the Summit Project within a 10,600 acre analysis area, located around and north of the community of Alta Sierra in the Greenhorn Mountains in the Kern River Ranger District of the Sequoia National Forest. The purpose of the project is “to provide defensible space around the community of Alta Sierra and nearby structures, improve forest health and provide for ecological restoration.” The Forest Service proposes to reduce fuels on 2,500 acres, or about 25% of the analysis area, through hand- and mechanical thinning so it can eventually re-introduce fire into the area. And while the Forest Service asserts that fuels reduction, ecological restoration and movement toward the desired conditions identified in the Sierra Nevada Forest Plan Amendment are the driving influences that will dictate the end result, it proposes to sell timber “as a tool to improve efficiency and reduce overall costs to achieve its fuel reduction and restoration goals.” The proposed thinning treatments may include commercial timber harvest within the 2,500 acre proposed treatment areas, which the Forest Service expects will produce roughly 5,000 CCF of timber sale volume.