Stock Status Report 2016
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ILLEGAL FISHING Which Fish Species Are at Highest Risk from Illegal and Unreported Fishing?
ILLEGAL FISHING Which fish species are at highest risk from illegal and unreported fishing? October 2015 CONTENTS EXECUTIVE SUMMARY 3 INTRODUCTION 4 METHODOLOGY 5 OVERALL FINDINGS 9 NOTES ON ESTIMATES OF IUU FISHING 13 Tunas 13 Sharks 14 The Mediterranean 14 US Imports 15 CONCLUSION 16 CITATIONS 17 OCEAN BASIN PROFILES APPENDIX 1: IUU Estimates for Species Groups and Ocean Regions APPENDIX 2: Estimates of IUU Risk for FAO Assessed Stocks APPENDIX 3: FAO Ocean Area Boundary Descriptions APPENDIX 4: 2014 U.S. Edible Imports of Wild-Caught Products APPENDIX 5: Overexploited Stocks Categorized as High Risk – U.S. Imported Products Possibly Derived from Stocks EXECUTIVE SUMMARY New analysis by World Wildlife Fund (WWF) finds that over 85 percent of global fish stocks can be considered at significant risk of Illegal, Unreported, and Unregulated (IUU) fishing. This evaluation is based on the most recent comprehensive estimates of IUU fishing and includes the worlds’ major commercial stocks or species groups, such as all those that are regularly assessed by the United Nations Food and Agriculture Organization (FAO). Based on WWF’s findings, the majority of the stocks, 54 percent, are categorized as at high risk of IUU, with an additional 32 perent judged to be at moderate risk. Of the 567 stocks that were assessed, the findings show that 485 stocks fall into these two categories. More than half of the world’s most overexploited stocks are at the highest risk of IUU fishing. Examining IUU risk by location, the WWF analysis shows that in more than one-third of the world’s ocean basins as designated by the FAO, all of these stocks were at high or moderate risk of IUU fishing. -

Otolith Trace Elemental Analyses of South American Austral Hake, Merluccius Australis
RESEARCH ARTICLE Otolith Trace Elemental Analyses of South American Austral Hake, Merluccius australis (Hutton, 1872) Indicates Complex Salinity Structuring on their Spawning/Larval Grounds Paul Brickle1,2,3*, Pia C. Schuchert4, Alexander I. Arkhipkin1, Malcolm R. Reid5, Haseeb S. Randhawa6 1 Directorate of Natural Resources, Fisheries Department, Falkland Islands Government, Stanley, Falkland Islands, 2 South Atlantic Environmental Research Institute, Stanley Cottage, Stanley, Falkland Islands, 3 School of Biological Sciences, University of Aberdeen, Zoology Building, Tillydrone Avenue, Aberdeen, AB24 2TZ, United Kingdom, 4 School of Biology, Newcastle University, Newcastle upon Tyne, NE1 7RU, United Kingdom, 5 Department of Chemistry, University of Otago, PO Box 56, Dunedin, 9054, New Zealand, 6 Ecology Degree Programme, Department of Botany, University of Otago, PO Box 56, Dunedin, 9054, New Zealand * [email protected] OPEN ACCESS Citation: Brickle P, Schuchert PC, Arkhipkin AI, Reid Abstract MR, Randhawa HS (2016) Otolith Trace Elemental Analyses of South American Austral Hake, Trace element signatures of otolith edges and cores from 335 austral hake (Merluccius Merluccius australis (Hutton, 1872) Indicates autralis) were analysed using LA-ICPMS from samples collected in Chilean and Falkland Complex Salinity Structuring on their Spawning/ Larval Grounds. PLoS ONE 11(1): e0145479. Islands' waters, in order to provide potential insights into stock discrimination and migra- doi:10.1371/journal.pone.0145479 tions. Fish were caught in two locations in Chile and four locations in the south-west of the Editor: Heather M. Patterson, Department of Falkland Islands Shelf. Univariate and multivariate analyses of trace element signatures in Agriculture and Water Resources, AUSTRALIA the edges of otoliths, representing adult fish, were not able to distinguish between samples ’ Received: July 27, 2014 collected in Chile and the Falkland Islands. -

Intrinsic Vulnerability in the Global Fish Catch
The following appendix accompanies the article Intrinsic vulnerability in the global fish catch William W. L. Cheung1,*, Reg Watson1, Telmo Morato1,2, Tony J. Pitcher1, Daniel Pauly1 1Fisheries Centre, The University of British Columbia, Aquatic Ecosystems Research Laboratory (AERL), 2202 Main Mall, Vancouver, British Columbia V6T 1Z4, Canada 2Departamento de Oceanografia e Pescas, Universidade dos Açores, 9901-862 Horta, Portugal *Email: [email protected] Marine Ecology Progress Series 333:1–12 (2007) Appendix 1. Intrinsic vulnerability index of fish taxa represented in the global catch, based on the Sea Around Us database (www.seaaroundus.org) Taxonomic Intrinsic level Taxon Common name vulnerability Family Pristidae Sawfishes 88 Squatinidae Angel sharks 80 Anarhichadidae Wolffishes 78 Carcharhinidae Requiem sharks 77 Sphyrnidae Hammerhead, bonnethead, scoophead shark 77 Macrouridae Grenadiers or rattails 75 Rajidae Skates 72 Alepocephalidae Slickheads 71 Lophiidae Goosefishes 70 Torpedinidae Electric rays 68 Belonidae Needlefishes 67 Emmelichthyidae Rovers 66 Nototheniidae Cod icefishes 65 Ophidiidae Cusk-eels 65 Trachichthyidae Slimeheads 64 Channichthyidae Crocodile icefishes 63 Myliobatidae Eagle and manta rays 63 Squalidae Dogfish sharks 62 Congridae Conger and garden eels 60 Serranidae Sea basses: groupers and fairy basslets 60 Exocoetidae Flyingfishes 59 Malacanthidae Tilefishes 58 Scorpaenidae Scorpionfishes or rockfishes 58 Polynemidae Threadfins 56 Triakidae Houndsharks 56 Istiophoridae Billfishes 55 Petromyzontidae -

A Review of Longnose Skates Zearaja Chilensisand Dipturus Trachyderma (Rajiformes: Rajidae)
Univ. Sci. 2015, Vol. 20 (3): 321-359 doi: 10.11144/Javeriana.SC20-3.arol Freely available on line REVIEW ARTICLE A review of longnose skates Zearaja chilensis and Dipturus trachyderma (Rajiformes: Rajidae) Carolina Vargas-Caro1 , Carlos Bustamante1, Julio Lamilla2 , Michael B. Bennett1 Abstract Longnose skates may have a high intrinsic vulnerability among fishes due to their large body size, slow growth rates and relatively low fecundity, and their exploitation as fisheries target-species places their populations under considerable pressure. These skates are found circumglobally in subtropical and temperate coastal waters. Although longnose skates have been recorded for over 150 years in South America, the ability to assess the status of these species is still compromised by critical knowledge gaps. Based on a review of 185 publications, a comparative synthesis of the biology and ecology was conducted on two commercially important elasmobranchs in South American waters, the yellownose skate Zearaja chilensis and the roughskin skate Dipturus trachyderma; in order to examine and compare their taxonomy, distribution, fisheries, feeding habitats, reproduction, growth and longevity. There has been a marked increase in the number of published studies for both species since 2000, and especially after 2005, although some research topics remain poorly understood. Considering the external morphological similarities of longnose skates, especially when juvenile, and the potential niche overlap in both, depth and latitude it is recommended that reproductive seasonality, connectivity and population structure be assessed to ensure their long-term sustainability. Keywords: conservation biology; fishery; roughskin skate; South America; yellownose skate Introduction Edited by Juan Carlos Salcedo-Reyes & Andrés Felipe Navia Global threats to sharks, skates and rays have been 1. -

S1755267211000431jra
S1755267211000431jra Author Queries No Queries Marine Biodiversity Records, page 1 of 3. # Marine Biological Association of the United Kingdom, 2011 doi:10.1017/S1755267211000431; Vol. 00; e0; 2011 Published online 1 2 First report of Macruronus novaezelandiae 3 4 (Gadiformes, Merluccidae, Macruroninae) 5 6 7 from Atlantic tropical waters 8 1 2 3 2 9 alfredo carvalho-filho , guy marcovaldi , cla’ udio l.s. sampaio and m. isabel g. paiva 10 1Fish-Bizz Ltda, Rua Maria Garcez, 39, Sa˜o Paulo, SP, 05424-070, Brazil, 2Projeto Tamar-ICMBio, Avenida do Farol Garcia D’A´ vila, 11 s/n, Praia do Forte, Mata de Sa˜o Joa˜o, BA, 48280-000, Brazil, 3Universidade Federal de Alagoas, Unidade de Ensino Penedo, Av. Beira 12 Rio s/n, Centro Histo´rico, Penedo, AL, 57.200-000, Brazil 13 14 15 The occurrence of the merluccid Macruronus novaezelandiae from tropical waters off Bahia, eastern Brazil, is reported for the 16 first time due to the capture of an adult of 712.3 mm SL in May 2008, from a depth of 400 metres. Until then no specimen had 17 been reported north of 32829′S on the South American Atlantic coast. This new record extends the species’ range to about 18 2500 km northwards along the Brazilian coastline and is the first ever from tropical waters in the world. A comparison of 19 the morphometric characters is provided. 20 21 22 Keywords: range extension, Macruronus magellanicus, deep-sea fish, Brazil 23 24 Submitted 11 December 2010; accepted 14 March 2011 25 26 27 INTRODUCTION also observed in several other species of the family belonging 28 to the genus Merluccius, already cited above. -

MERLUZA AUSTRAL (Merluccius Australis)
MERLUZA AUSTRAL (Merluccius australis) por Analía R. Giussi, Susana B. García de la Rosa y Felisa Sánchez IDENTIFICACIÓN DEL RECURSO Clase: Actinopterygii. Orden: Gadiformes. Familia: Merlucciidae. Especie: Merluccius australis (Hutton, 1872). Nombre común:merluza austral, merluzón (Argentina); merluza del sur (Chile). Nombre en inglés: southern hake. Otros nombres científicos sinónimos en uso: Merluccius polylepis y Merluccius australis polylepis. DISTRIBUCIÓN GEOGRÁFICA La merluza austral es una especie que se halla ampliamente distribuida en el hemisferio sur, tanto en aguas argentinas y chilenas como en neozelandesas (Cousseau y Perrotta, 1998). En el extremo sur de América ocupa un área que se extiende, en el Océano Pacífico, al sur de los 40°S entre 50 y 600 m de pro- fundidad (Aguayo-Hernández, 1994), y en el Océano Atlántico al sur de los 50°S, desde los 100 a los 400 m de profundidad (García de la Rosa et al., 1997). Esta distribución es continua a través del Pasaje de Drake (Aguayo, 1995), hallándose individuos en la región norte del Estrecho de Magallanes (Céspedes et al., 1996). Esta especie se caracteriza por presentar hábitos demersales y está relacionada, en el Mar Argentino, con aguas frías de la Corriente de Malvinas (Otero y Simonazzi, 1980), localizándose sus ma- yores concentraciones entre 50°-55°S (García de la Rosa et al., 1997) (Figura 1). García de la Rosa et al. (1997), analizando la distribución de esta especie en el transcurso del año, observaron que en el verano las mayores concentraciones (9 y 13 t/mn2) se hallaron al sur de la Isla de los Estados (54°50'S), extendiéndose su área de distribución desde los 48° hasta 55°S y entre 100 y 200 m de profundidad. -

Biology and Fishery of Long Tail Hake (Macruronus Magellanicus) in the Southwest Atlantic Ocean
Biology and fishery of long tail hake (Macruronus magellanicus) in the Southwest Atlantic Ocean. Item Type Journal Contribution Authors Giussi, A.R.; Zavatteri, A.; Di Marco, E.J.; Gorini, F.L.; Bernardele, J.C.; Marí, N. Download date 03/10/2021 00:46:44 Link to Item http://hdl.handle.net/1834/10967 REV. INVEST. DESARR. PESQ. Nº 28: 55-82 (2016) 55 REVISIÓN BIOLOGY AND FISHERY OF LONG TAIL HAKE (Macruronus magellanicus) IN THE SOUTHWEST ATLANTIC OCEAN* ANALÍA R. GIUSSI1, ANABELA ZAVATTERI, EMILIANO J. DI MARCO, FEDERICO L. GORINI, JUAN C. BERNARDELE† and NOEMÍ R. MARÍ Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP), Paseo Victoria Ocampo N° 1, Escollera Norte, B7602HSA - Mar del Plata, Argentina 1e-mail: [email protected] SUMMARY. Long tail hake is one of the most important finfish resources in the Southwest Atlantic Ocean (SAO). This demersal-pelagic fish is widely distributed from 35° S to 56° S between 50 and 800 m depth. In the Patagonian region the species is found on the shelf, associated with three different water masses: Coastal, Shelf and Malvinas Waters, while north of 45° S it is related to the shelf break, following the Malvinas waters. A separate small stock, evident because of its different growth patterns, occurs inside San Matías Gulf. There is no strong difference in growth patterns between sexes, however females are larger than males, principally after the first maturity that occurs at 3 years old. Maximum age observed was 16 years old, but fish older than 12 years are scarce in the population. The dietary composition changes during the life cycle, juveniles are mainly microphagous and adults incorporate larger preys of several invertebrates and vertebrates taxa. -

Fish Stocks United Nations Food and Agriculture Organization (FAO)
General situation of world fish stocks United Nations Food and Agriculture Organization (FAO) Contents: 1. Definitions 2. Snapshot of the global situation 3. Short list of "depleted" fish stocks 4. Global list of fish stocks ranked as either "overexploited," "depleted," or recovering by region 1. Definitions Underexploited Undeveloped or new fishery. Believed to have a significant potential for expansion in total production; Moderately exploited Exploited with a low level of fishing effort. Believed to have some limited potential for expansion in total production; Fully exploited The fishery is operating at or close to an optimal yield level, with no expected room for further expansion; Overexploited The fishery is being exploited at above a level which is believed to be sustainable in the long term, with no potential room for further expansion and a higher risk of stock depletion/collapse; Depleted Catches are well below historical levels, irrespective of the amount of fishing effort exerted; Recovering Catches are again increasing after having been depleted 2. Snapshot of the global situation Of the 600 marine fish stocks monitored by FAO: 3% are underexploited 20% are moderately exploited 52% are fully exploited 17% are overexploited 7% are depleted 1% are recovering from depletion Map of world fishing statistical areas monitored by FAO Source: FAO's report "Review of the State of World Marine Fisheries Resources", tables D1-D17, ftp://ftp.fao.org/docrep/fao/007/y5852e/Y5852E23.pdf 3. Fish stocks identified by FAO as falling into its -

Description of Merluccius Tasmanicus Sp. Nov. and Redescription
J. Mar. Biol. Ass. U.K. (2006), 86,193^199 Printed in the United Kingdom Description of Merluccius tasmanicus sp.nov.and redescription of Merluccius australis (Pisces: Merlucciidae) P O J. Matallanas* and D. Lloris *Unidad de Zoolog|¤a, Departamento de Biolog|¤a Animal, Biolog|¤aVegetal y Ecolog|¤a. Universidad Auto¤noma de Barcelona, 08193, O Bellaterra, Barcelona, Spain. Institut de Cie' ncies del Mar (CMIMA-CSIC), Passeig Mar|¤tim de la Barceloneta 37^49, P 08003 Barcelona, Spain. E-mail: [email protected]. Corresponding author, e-mail: [email protected] A new hake species, Merluccius tasmanicus sp. nov., is described from New Zealand waters and another species, Merluccius australis is redescribed. Merluccius tasmanicus sp. nov. di¡ers from all other congeneric species in the following combination of characters: upper pro¢le of the head slowly concave; lateral line slowly concave in the caudal region; body depth 4.9^5.9 times in standard length (SL); orbital diameter 6.1^7.1 times in head length, 2.1^2.2 times in snout length and 1.6^1.9 times in interorbital width; second dorsal ¢n rays, 42^43; anal ¢n rays, 42^44; lateral line scales *164. Merluccius australis is redescribed to clarify the identity of this species. Merluccius australis di¡ers from all other congeneric species in the following combination of characters: upper pro¢le of the head straight; lateral line straight in the caudal region; body depth 6.6^7.1times in SL; orbital diameter 4.5^5.4 times in head length, 1.2^1.7 times in snout length and 1.0^1.3 times in interorbital width; second dorsal ¢n rays, 40^43; anal ¢n rays, 40^43; lateral line scales, more than 155. -

Professores - Produção Científica
PROFESSORES - PRODUÇÃO CIENTÍFICA PROF. DR. ALDO RAMOS SANTOS Artigos completos publicados em periódicos 1. CESAR, A ; Denis Moledo Abessa ; PEREIRA, C. D. S. ; SANTOS, A. R. Simple Approach to Integrate Ecotoxicological and Chemical Data for the Establishment of Environmental Risk Levels.. Brazilian Archives of Biology and Technology , v. v. 52, p. 233-240, 2009. 2. SANTOS, A. R. ; SILVA, Rogério José da . Análisis del Consumo de Coque de Petroleo en Algunos Sectores Industriales. Información Tecnológica , v. 1, p. 93-101, 2008. 3. SANTOS, A. R. ; SILVA, Rogério José da . Comparação entre o processamento de petróleos convencionais e não convencionais e os impactos causados no segmento de refino. Revista Ceciliana (Santos) , v. Ano 19, p. 85-104, 2008. 4. CESAR, A ; Denis Moledo Abessa ; PEREIRA, C. D. S. ; SANTOS, A. R. ; Rodrigo Brasil Choueri ; Nuria Fernández ; Tomaz Angel Del Valls Casillas . Comparative sediment quality assessment in different littoral ecossystems from Spain (Gulf of Cádiz) and Brazil (Santos and São Vicente estuarine systm). Environment International , v. 33, p. 429-435, 2007. 5. CESAR, A ; PEREIRA, C. D. S. ; SANTOS, A. R. ; Denis Moledo Abessa ; Nuria Fernández ; Rodrigo Brasil Choueri ; Tomaz Angel Del Valls Casillas . Ecotoxicology assessment of sediments from Santos and São Vicente Estuarine System Brazil. Brazilian Journal of Oceanography (Impresso) , v. 54, p. 55-63, 2006. PROF. DR. ALVARO LUIZ DIOGO REIGADA Artigos completos publicados em periódicos 1. SANT`ANNA, Bruno Sampaio ; REIGADA, A. L. D. ; PINHEIRO, Marcelo Antonio Amaro . Population biology and reproduction of the hermit crab Clibanarius vittatus (Decapoda: Anomura) in an estuarine region of southern Brazil. Journal of the Marine Biological Association of the United Kingdom , v. -

Stock Assessment of Hake (Merluccius Australis) on Chatham Rise for the 2019–20 Fishing Year
Stock assessment of hake (Merluccius australis) on Chatham Rise for the 2019–20 fishing year New Zealand Fisheries Assessment Report 2021/22 S.J. Holmes ISSN 1179-5352 (online) ISBN 978-1-99-100374-4 (online) April 2021 Requests for further copies should be directed to: Publications Logistics Officer Ministry for Primary Industries PO Box 2526 WELLINGTON 6140 Email: [email protected] Telephone: 0800 00 83 33 Facsimile: 04-894 0300 This publication is also available on the Ministry for Primary Industries websites at: http://www.mpi.govt.nz/news-and-resources/publications http://fs.fish.govt.nz go to Document library/Research reports © Crown Copyright – Fisheries New Zealand TABLE OF CONTENTS EXECUTIVE SUMMARY 1 1. INTRODUCTION 2 1.1 Description of the fishery 2 1.2 Literature review 3 2. BIOLOGY, STOCK STRUCTURE, AND ABUNDANCE INDICES 4 2.1 Biology 4 2.2 Stock structure 6 2.3 Resource surveys 6 2.4 Observer age samples 7 2.5 CPUE indices 8 3. MODEL STRUCTURE, INPUTS, AND ESTIMATION 9 3.1 Prior distributions and penalty functions 14 4. MODEL ESTIMATES FOR CHATHAM RISE HAKE 14 4.1 Developing a ‘base’ model 14 4.2 Model estimation using MCMC 21 4.3 Biomass projections 26 4.4 Management biomass targets 29 5. DISCUSSION 30 6. ACKNOWLEDGMENTS 31 7. REFERENCES 32 APPENDIX A: RESOURCE SURVEY BIOMASS INDICES FOR HAKE IN HAK 4 36 APPENDIX B: MPD FITS TO COMPOSITION DATA 38 APPENDIX C: MCMC CONVERGENCE AND DISTRIBUTION PLOTS 43 EXECUTIVE SUMMARY Holmes, S.J. (2021). Stock assessment of hake (Merluccius australis) on Chatham Rise for the 2019–20 fishing year. -
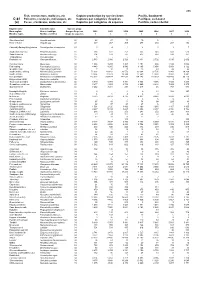
Fish, Crustaceans, Molluscs, Etc Capture Production by Species
495 Fish, crustaceans, molluscs, etc Capture production by species items Pacific, Southwest C-81 Poissons, crustacés, mollusques, etc Captures par catégories d'espèces Pacifique, sud-ouest (a) Peces, crustáceos, moluscos, etc Capturas por categorías de especies Pacífico, sudoccidental English name Scientific name Species group Nom anglais Nom scientifique Groupe d'espèces 2002 2003 2004 2005 2006 2007 2008 Nombre inglés Nombre científico Grupo de especies t t t t t t t Short-finned eel Anguilla australis 22 28 27 13 10 5 ... ... River eels nei Anguilla spp 22 337 267 209 277 210 207 152 Chinook(=Spring=King)salmon Oncorhynchus tshawytscha 23 0 4 1 2 1 1 7 Southern lemon sole Pelotretis flavilatus 31 238 322 251 335 348 608 513 Sand flounders Rhombosolea spp 31 204 193 187 437 514 530 351 Tonguefishes Cynoglossidae 31 3 - - - - - - Flatfishes nei Pleuronectiformes 31 2 580 2 986 2 729 3 431 2 702 3 015 2 602 Common mora Mora moro 32 1 308 1 234 1 403 1 154 986 1 180 1 088 Red codling Pseudophycis bachus 32 4 443 8 265 9 540 8 165 5 854 5 854 6 122 Grenadier cod Tripterophycis gilchristi 32 7 10 13 13 43 29 26 Southern blue whiting Micromesistius australis 32 72 203 43 812 26 576 30 304 32 735 23 943 29 268 Southern hake Merluccius australis 32 13 834 22 623 19 344 12 560 12 858 13 892 8 881 Blue grenadier Macruronus novaezelandiae 32 215 302 209 414 147 032 134 145 119 329 103 489 96 119 Ridge scaled rattail Macrourus carinatus 32 - - - - - 9 14 Thorntooth grenadier Lepidorhynchus denticulatus 32 5 349 5 304 6 341 3 855 4 056 3 725 3 264 Grenadiers, rattails nei Macrouridae 32 3 877 4 253 3 732 2 660 2 848 7 939 8 970 Gadiformes nei Gadiformes 32 3 252 3 281 298 1 217 46 767 886 Broadgilled hagfish Eptatretus cirrhatus 33 2 - 0 0 11 508 347 Sea catfishes nei Ariidae 33 4 6 4 4 4 ..