Acacia, Cattle and Migratory Birds in Southeastern Mexico
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Assessing Bird Species Richness Within Shade-Grown Coffee Farms in Chiapas, Mexico / Project ID: 0251711
Assessing Bird Species Richness within Shade-Grown Coffee Farms in Chiapas, Mexico / Project ID: 0251711 Daniel Camilo Thompson Poo, Daniela Valle León, Alberto Martínez Fernández and Jennifer Siobhan Lowry San Cristóbal de las Casas, Chiapas, México. C.P. 29200 / [email protected] 10 July, 2012. Revised December 2014 Assessing Bird Species Richness within Shade-Grown Coffee Farms in Chiapas, Mexico / ID: 0251711 Overall Aim The goal of this project was to identify mechanisms and conservation strategies across agro-forestry systems in the El Triunfo Biosphere Reserve in Chiapas, Mexico. In particular we analyzed key biodiversity, economic, and social components that impact land-use change and ecosystem services in coffee production areas, focusing on how to improve sustainable production and conservation of nature. 2 Assessing Bird Species Richness within Shade-Grown Coffee Farms in Chiapas, Mexico / ID: 0251711 Section 1 Summary The agroforestry systems with coffee at the Sierra Madre of Chiapas, as a part of the Mesoamerican Biological Corridor region, are important for bird species. Agroforestry ecosystems also represent sustainable livelihoods for indigenous groups on the region. Sustainable coffee farming system represents a less human impact on the ecosystem. However, not all coffee producers on the region produce on the same way. Not all the inhabitants are aware of the importance of birds, as a part of the great natural capital of la Sierra Madre, but they either are prepared for the climate change risks and impacts. In this sense, this project seeks to understand, generate and communicate information useful for coffee farmers and their families. The goal is to understand social and economic factors to maintain and increase agroforestry systems with sustainable coffee. -

Tinamiformes – Falconiformes
LIST OF THE 2,008 BIRD SPECIES (WITH SCIENTIFIC AND ENGLISH NAMES) KNOWN FROM THE A.O.U. CHECK-LIST AREA. Notes: "(A)" = accidental/casualin A.O.U. area; "(H)" -- recordedin A.O.U. area only from Hawaii; "(I)" = introducedinto A.O.U. area; "(N)" = has not bred in A.O.U. area but occursregularly as nonbreedingvisitor; "?" precedingname = extinct. TINAMIFORMES TINAMIDAE Tinamus major Great Tinamou. Nothocercusbonapartei Highland Tinamou. Crypturellus soui Little Tinamou. Crypturelluscinnamomeus Thicket Tinamou. Crypturellusboucardi Slaty-breastedTinamou. Crypturellus kerriae Choco Tinamou. GAVIIFORMES GAVIIDAE Gavia stellata Red-throated Loon. Gavia arctica Arctic Loon. Gavia pacifica Pacific Loon. Gavia immer Common Loon. Gavia adamsii Yellow-billed Loon. PODICIPEDIFORMES PODICIPEDIDAE Tachybaptusdominicus Least Grebe. Podilymbuspodiceps Pied-billed Grebe. ?Podilymbusgigas Atitlan Grebe. Podicepsauritus Horned Grebe. Podicepsgrisegena Red-neckedGrebe. Podicepsnigricollis Eared Grebe. Aechmophorusoccidentalis Western Grebe. Aechmophorusclarkii Clark's Grebe. PROCELLARIIFORMES DIOMEDEIDAE Thalassarchechlororhynchos Yellow-nosed Albatross. (A) Thalassarchecauta Shy Albatross.(A) Thalassarchemelanophris Black-browed Albatross. (A) Phoebetriapalpebrata Light-mantled Albatross. (A) Diomedea exulans WanderingAlbatross. (A) Phoebastriaimmutabilis Laysan Albatross. Phoebastrianigripes Black-lootedAlbatross. Phoebastriaalbatrus Short-tailedAlbatross. (N) PROCELLARIIDAE Fulmarus glacialis Northern Fulmar. Pterodroma neglecta KermadecPetrel. (A) Pterodroma -

21 Sep 2018 Lists of Victims and Hosts of the Parasitic
version: 21 Sep 2018 Lists of victims and hosts of the parasitic cowbirds (Molothrus). Peter E. Lowther, Field Museum Brood parasitism is an awkward term to describe an interaction between two species in which, as in predator-prey relationships, one species gains at the expense of the other. Brood parasites "prey" upon parental care. Victimized species usually have reduced breeding success, partly because of the additional cost of caring for alien eggs and young, and partly because of the behavior of brood parasites (both adults and young) which may directly and adversely affect the survival of the victim's own eggs or young. About 1% of all bird species, among 7 families, are brood parasites. The 5 species of brood parasitic “cowbirds” are currently all treated as members of the genus Molothrus. Host selection is an active process. Not all species co-occurring with brood parasites are equally likely to be selected nor are they of equal quality as hosts. Rather, to varying degrees, brood parasites are specialized for certain categories of hosts. Brood parasites may rely on a single host species to rear their young or may distribute their eggs among many species, seemingly without regard to any characteristics of potential hosts. Lists of species are not the best means to describe interactions between a brood parasitic species and its hosts. Such lists do not necessarily reflect the taxonomy used by the brood parasites themselves nor do they accurately reflect the complex interactions within bird communities (see Ortega 1998: 183-184). Host lists do, however, offer some insight into the process of host selection and do emphasize the wide variety of features than can impact on host selection. -

Engelsk Register
Danske navne på alverdens FUGLE ENGELSK REGISTER 1 Bearbejdning af paginering og sortering af registret er foretaget ved hjælp af Microsoft Excel, hvor det har været nødvendigt at indlede sidehenvisningerne med et bogstav og eventuelt 0 for siderne 1 til 99. Tallet efter bindestregen giver artens rækkefølge på siden. -

The Brown Jay's Furcular Pouch
160 Vol. 44 THE BROWN JAY’S FURCULAR POUCH By GEORGE MIKSCH SUTTON and PERRY W. GILBERT Travelers along northeastern Mexico’s modern highway are apt to remark upon the scarcity of bird life. Those who motor at reasonable speed from Monterrey to Victoria, and thence to Mante, Valles, and Tamazunchale, are sure to see, and almost sure to hear, however, a crow-sized, dark gray-brown bird, with long&h tail, that in- habits the brush of the low country. This bird is in no way handsome, but its boldness, simple coloration, and obvious predilection for the roadside insure its being noticed and talked about. It belongs to that tribe of mischief makers, the Corvidae. It is the Brown Jay, Psilorhinus morio. Fig. 33. Brown Jay (Psilorhinus morio), about one-fiith iife size. Drawing by George Miksch Sutton. July, 1942 THE BROWN JAY’S FURCULAR POUCH 161 The Brown Jay almost never goes about alone. Family groups of ,five or six are the rule, although sometimes when an owl, lynx, human being, or other enemy species is being mobbed, a party of twenty or more may congregate. Excitement then runs high. Each jay must approach ‘Yhe enemy” as closely as it can, so as personally to confirm the “rumors” it has heard. Each jay must scream. If “the enemy” moves, the outcry momentarily stops short; the jays eye each other as if pondering the next move; and the hubbub of screaming starts afresh. When the senior author first encountered the Brown Jay in the vicinity of Monterrey, Nuevo Leon, in February of 1938, he was greatly struck with what he called the bird’s ‘Lhiccup.” This sound, which usually was heard just at the close of the customary scream, but often quite independently of it, was not unlike the syllable puck or buck. -

Gear for a Big Year
APPENDIX 1 GEAR FOR A BIG YEAR 40-liter REI Vagabond Tour 40 Two passports Travel Pack Wallet Tumi luggage tag Two notebooks Leica 10x42 Ultravid HD-Plus Two Sharpie pens binoculars Oakley sunglasses Leica 65 mm Televid spotting scope with tripod Fossil watch Leica V-Lux camera Asics GEL-Enduro 7 trail running shoes GoPro Hero3 video camera with selfie stick Four Mountain Hardwear Wicked Lite short-sleeved T-shirts 11” MacBook Air laptop Columbia Sportswear rain shell iPhone 6 (and iPhone 4) with an international phone plan Marmot down jacket iPod nano and headphones Two pairs of ExOfficio field pants SureFire Fury LED flashlight Three pairs of ExOfficio Give- with rechargeable batteries N-Go boxer underwear Green laser pointer Two long-sleeved ExOfficio BugsAway insect-repelling Yalumi LED headlamp shirts with sun protection Sea to Summit silk sleeping bag Two pairs of SmartWool socks liner Two pairs of cotton Balega socks Set of adapter plugs for the world Birding Without Borders_F.indd 264 7/14/17 10:49 AM Gear for a Big Year • 265 Wildy Adventure anti-leech Antimalarial pills socks First-aid kit Two bandanas Assorted toiletries (comb, Plain black baseball cap lip balm, eye drops, toenail clippers, tweezers, toothbrush, REI Campware spoon toothpaste, floss, aspirin, Israeli water-purification tablets Imodium, sunscreen) Birding Without Borders_F.indd 265 7/14/17 10:49 AM APPENDIX 2 BIG YEAR SNAPSHOT New Unique per per % % Country Days Total New Unique Day Day New Unique Antarctica / Falklands 8 54 54 30 7 4 100% 56% Argentina 12 435 -

Section IV – Guideline for the Texas Priority Species List
Section IV – Guideline for the Texas Priority Species List Associated Tables The Texas Priority Species List……………..733 Introduction For many years the management and conservation of wildlife species has focused on the individual animal or population of interest. Many times, directing research and conservation plans toward individual species also benefits incidental species; sometimes entire ecosystems. Unfortunately, there are times when highly focused research and conservation of particular species can also harm peripheral species and their habitats. Management that is focused on entire habitats or communities would decrease the possibility of harming those incidental species or their habitats. A holistic management approach would potentially allow species within a community to take care of themselves (Savory 1988); however, the study of particular species of concern is still necessary due to the smaller scale at which individuals are studied. Until we understand all of the parts that make up the whole can we then focus more on the habitat management approach to conservation. Species Conservation In terms of species diversity, Texas is considered the second most diverse state in the Union. Texas has the highest number of bird and reptile taxon and is second in number of plants and mammals in the United States (NatureServe 2002). There have been over 600 species of bird that have been identified within the borders of Texas and 184 known species of mammal, including marine species that inhabit Texas’ coastal waters (Schmidly 2004). It is estimated that approximately 29,000 species of insect in Texas take up residence in every conceivable habitat, including rocky outcroppings, pitcher plant bogs, and on individual species of plants (Riley in publication). -

The Puzzling Vocal Repertoire of the South American Collared Jay, Cyanolyca Iridicyana Merida
THE PUZZLING VOCAL REPERTOIRE OF THE SOUTH AMERICAN COLLARED JAY, CYANOLYCA VZRIDZCYANA MERIDA JOHN WILLIAM HARDY Previously, I have indicated (Kansas Sci. Bull., 42: 113, 1961) that the New World jays could be systematically grouped in two tribes on the basis of a variety of morphological and behavioral characteristics. I subsequently demonstrated (Occas. Papers Adams Ctr. Ecol. Studies, No. 11, 1964) the relationships between Cyanolyca and Aphelocoma. The tribe Aphelocomini, comprising the currently recognized genera Aphelocoma and Cyanolyca, may be characterized as follows: pattern and ornamen- tation of head simple, consisting of a dark mask, pale superciliary lineation, and no tendency for a crest; tail plain-tipped; vocal repertoire of one to three basic com- ponents, including alarm calls, flock-social calls, or both, that are nasal, querulous, and upwardly or doubly inflected. In contrast, the Cyanocoracini may be charac- tized as follows: pattern of head complex, consisting of triangular cheek patch, super- ciliary spot, and tendency for a crest; tail usually pale-tipped; vocal repertoire usu- ally complex, commonly including a downwardly inflected cuwing call and never an upwardly inflected, nasal querulous call (when repertoire is limited it almost always includes the cazo!ingcall). None of the four Mexican aphelocomine jays has more than three basic calls in its vocal repertoire. There are two exclusively South American species of CyanuZyca (C. viridicyana and C. pulchra). On the basis of the consistency with which the Mexican and Central American speciesof this genus hew to a general pattern of vocalizations as I described it for the tribe, I inferred (1964 op. -

The White-Throated Magpie-Jay
THE WHITE-THROATED MAGPIE-JAY BY ALEXANDER F. SKUTCH ORE than 15 years have passed since I was last in the haunts of the M White-throated Magpie-Jay (Calocitta formosa) . During the years when I travelled more widely through Central America I saw much of this bird, and learned enough of its habits to convince me that it would well repay a thorough study. Since it now appears unlikely that I shall make this study myself, I wish to put on record such information as I gleaned, in the hope that these fragmentary notes will stimulate some other bird- watcher to give this jay the attention it deserves. A big, long-tailed bird about 20 inches in length, with blue and white plumage and a high, loosely waving crest of recurved black feathers, the White-throated Magpie-Jay is a handsome species unlikely to be confused with any other member of the family. Its upper parts, including the wings and most of the tail, are blue or blue-gray with a tinge of lavender. The sides of the head and all the under plumage are white, and the outer feathers of the strongly graduated tail are white on the terminal half. A narrow black collar crosses the breast and extends half-way up each side of the neck, between the white and the blue. The stout bill and the legs and feet are black. The sexes are alike in appearance. The species extends from the Mexican states of Colima and Puebla to northwestern Costa Rica. A bird of the drier regions, it is found chiefly along the Pacific coast from Mexico southward as far as the Gulf of Nicoya in Costa Rica. -

Distribution Habitat and Social Organization of the Florida Scrub Jay
DISTRIBUTION, HABITAT, AND SOCIAL ORGANIZATION OF THE FLORIDA SCRUB JAY, WITH A DISCUSSION OF THE EVOLUTION OF COOPERATIVE BREEDING IN NEW WORLD JAYS BY JEFFREY A. COX A DISSERTATION PRESENTED TO THE GRADUATE COUNCIL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 1984 To my wife, Cristy Ann ACKNOWLEDGEMENTS I thank, the following people who provided information on the distribution of Florida Scrub Jays or other forms of assistance: K, Alvarez, M. Allen, P. C. Anderson, T. H. Below, C. W. Biggs, B. B. Black, M. C. Bowman, R. J. Brady, D. R. Breininger, G. Bretz, D. Brigham, P. Brodkorb, J. Brooks, M. Brothers, R. Brown, M. R. Browning, S. Burr, B. S. Burton, P. Carrington, K. Cars tens, S. L. Cawley, Mrs. T. S. Christensen, E. S. Clark, J. L. Clutts, A. Conner, W. Courser, J. Cox, R. Crawford, H. Gruickshank, E, Cutlip, J. Cutlip, R. Delotelle, M. DeRonde, C. Dickard, W. and H. Dowling, T. Engstrom, S. B. Fickett, J. W. Fitzpatrick, K. Forrest, D. Freeman, D. D. Fulghura, K. L. Garrett, G. Geanangel, W. George, T. Gilbert, D. Goodwin, J. Greene, S. A. Grimes, W. Hale, F. Hames, J. Hanvey, F. W. Harden, J. W. Hardy, G. B. Hemingway, Jr., 0. Hewitt, B. Humphreys, A. D. Jacobberger, A. F. Johnson, J. B. Johnson, H. W. Kale II, L. Kiff, J. N. Layne, R. Lee, R. Loftin, F. E. Lohrer, J. Loughlin, the late C. R. Mason, J. McKinlay, J. R. Miller, R. R. Miller, B. -
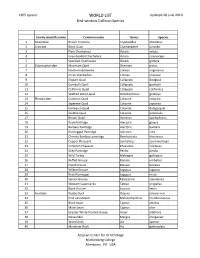
WORLD LIST Updated 28 June 2019 Bird-Window Collision Species
1305 species WORLD LIST Updated 28 June 2019 Bird-window Collision Species Family scientific name Common name Genus Species 1 Tinamidae Brown Tinamou Crypturellus obsoletus 2 Cracidae Black Guan Chamaepetes unicolor 3 Plain Chachalaca Ortalis vetula 4 Grey-headed Chachalaca Ortalis cinereiceps 5 Speckled Chachalaca Ortalis guttata 6 Odontophoridae Mountain Quail Oreortyx pictus 7 Northern Bobwhite Colinus virginianus 8 Crested Bobwhite Colinus cristatus 9 Elegant Quail Callipepla douglasii 10 Gambel's Quail Callipepla gambelii 11 California Quail Callipepla californica 12 Spotted Wood-quail Odontophorus guttatus 13 Phasianidae Common Quail Coturnix coturnix 14 Japanese Quail Coturnix japonica 15 Harlequin Quail Coturnix delegorguei 16 Stubble Quail Coturnix pectoralis 17 Brown Quail Synoicus ypsilophorus 18 Rock Partridge Alectoris graeca 19 Barbary Partridge Alectoris barbara 20 Red-legged Partridge Alectoris rufa 21 Chinese Bamboo-partridge Bambusicola thoracicus 22 Copper Pheasant Syrmaticus soemmerringii 23 Common Pheasant Phasianus colchicus 24 Grey Partridge Perdix perdix 25 Wild Turkey Meleagris gallopavo 26 Ruffed Grouse Bonasa umbellus 27 Hazel Grouse Bonasa bonasia 28 Willow Grouse Lagopus lagopus 29 Rock Ptarmigan Lagopus muta 30 Spruce Grouse Falcipennis canadensis 31 Western Capercaillie Tetrao urogallus 32 Black Grouse Lyrurus tetrix 33 Anatidae Ruddy Duck Oxyura jamaicensis 34 Pink-eared Duck Malacorhynchus membranaceus 35 Black Swan Cygnus atratus 36 Mute Swan Cygnus olor 37 Greater White-fronted Goose Anser albifrons 38 -

Southern Belize: Birding & Nature with Naturalist Journeys & Caligo Ventures
Southern Belize: Birding & Nature With Naturalist Journeys & Caligo Ventures March 10 – 18, 2018 With Crooked Tree Wildlife Sanctuary Extension Mar. 8 – 10 866.900.1146 800.426.7781 520.558.1146 [email protected] www.naturalistjourneys.com or find us on Facebook at Naturalist Journeys, LLC Naturalist Journeys, LLC / Caligo Ventures PO Box 16545 Portal, AZ 85632 PH: 520.558.1146 / 800.426.7781 Fax 650.471.7667www.naturalistjourneys.com / www.caligo.com [email protected] / [email protected] Southern Belize: Birding & Nature With Naturalist Journeys & Caligo Ventures This Belize birding tour explores the enchanting blend of Mayan ruins, exquisite coastline, and lush tropical forests that host jaguars, tapirs, and an exciting array of tropical birds, and lures visitors to Belize, Central America’s least populated country. The welcoming Belizean people speak English (often with Spanish and Mayan as second languages), have a strong tradition of caretaking their country’s natural heritage, and enjoy sharing their knowledge of the region’s biodiversity. Some of the very best wildlife and birding guides in Latin America live and work in Belize. Our Belize tour traverses several of the country’s distinctive ecoregions, focusing on the submontane Caribbean pine habitat and waterfalls of the rugged Maya Mountains, as well as the lowland tropical pine and broad-leaved forests and open savannas of the Cayo and Toledo Districts near the Caracol archaeological site and the villages of Big Falls and Red Bank. We also spend a day visiting tropical riparian scrub, mangrove, and seagrass environments along the Monkey River north of Punta Gorda. We stay four nights each at two outstanding lodges, allowing us time to settle in at our locations while reducing the number of hours spent on the road.