Genetic Polymorphism and Evolution in Parthenogenetic Animals
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vereine Kanton Glarus
Telefon 055 646 62 46 E-Mail: [email protected] www.gl.ch Bildung und Kultur Integration Stand: 11.06.2021 Gerichtshausstrasse 25 8750 Glarus Vereine Kanton Glarus Sport/Freizeitaktivitäten Verein / Anbieter Adresse Kontaktperson Ort Tel. / E-Mail Beschrieb Aikido Glarus Zinggenstrasse 2 Hansjörg Weber Mollis 078 660 05 66 Japanische Kampfkunst Badminton Club Glarus Platte 28 Dirk Sewing Ennenda 055 640 69 53 Badminton spielen Blauring Näfels Autschachen 1 Fränzi Fischli Näfels 079 373 77 66 Verschiedene Niederurnen Fronalpstrasse 21 Sibylle Bodenmann Niederurnen 055 610 18 64 Freizeitbetätigungen für Kinder. Christlicher Glaube. Seite 1 Fussballclub Glarus Adlerguet 19a Renato Micheroli Glarus 055 640 88 17 Fussball Fussballclub Linth 04 Erich Fischli Näfels 079 693 53 22 Fussballclub Netstal Schlöffeli 16 Moor Raphaël Netstal 079 244 10 11 Fussballclub Rüti GL Dorfstrasse 90 Roger Nievergelt Rüti 079 332 76 01 Fussballclub Schwanden Bahnhofstrasse 31 René Schätti Schwanden 079 575 31 61 Rythmische Gymnastik Speerstrasse 13 Stephanie Blunschi Näfels 079 596 94 12 Turnen & Gymnastik https://www.gltv.ch/rg Glaronia Volleyballclub Lurigenstrasse 17 Peter Aebli Glarus 055 640 37 24 Volleyball Judoclub Yawara Glarnerland Fabrikstrasse 3 Claudio Nicoletti Niederurnen 055 640 31 35 Judo Seite 2 Karateschule Glarus Landstrasse 41 / Daniel Rimann Glarus 055 650 19 30 Karate / Kickboxen Zaunturnhalle 079 421 14 06 KiTu-Kinderturnen Hätzingen- Turnhalle Hätzingen Elsbeth Mächler Hätzingen 079 695 87 66 Turnen / Gymnastik Luchsingen Leichtathletikverein Glarus Bifang 1 Reto Menzi Filzbach 079 787 80 45 Turnen / Gymnastik www.lavglarus.ch MuKi Hätzingen-Luchsingen Sonnenhof Liz Schindler Leuggelbach 076 436 2313 Turnen für Mütter/Väter mit MuKi-Turnen Bilten Hauptstrasse 58a Ruth Marti Bilten 055 610 21 54 Kind von 3-5 J. -

Behörden Der Kirchen
BEHÖRDEN DER KIRCHEN Evangelisch-Reformierte Landeskirche Synode Büro Präsident Hefti Andreas, lic. iur., Glarus Vizepräsidentin Lienhard Marianne, Landesstatthalter, Elm Aktuar Paysen-Petersen Jacqueline, Rufi 1. Stimmenzähler Hefti Hansheinrich, Schwanden 2. Stimmenzäler Wachsmuth Michael, Mitlödi Im Amt stehende Pfarrer und Pfarrerinnen Jahrgang Amtsantritt Neumann Almut, Mitlödi (Provisorin) 1952 1992 Rhyner-Funk Andrea, Elm (Pfarramt für Behinderte) 1965 2006 Brüll Beck Christina, Mollis 1972 2008 Doll Dagmar, Glarus 1973 2009 Doll Sebastian, Glarus 1972 2009 Hofmann Peter, Ennenda 1966 2011 Lustenberger Iris, Ennenda 1967 2011 Peters Matthias, Niederurnen 1958 2011 Zubler Daniel (Spitalpfarramt), Glarus 1961 2012 Aerni Edi, Netstal 1959 2014 Schneider Christoph, Betschwanden 1968 2015 Wüthrich Beat Emanuel, Matt 1958 2016 Pfiffner Annemarie, Obstalden 1963 2017 Wyler-Eschle Bruno, Bilten (Provisor) Von den Kirchgemeinden gewählte 50 Abgeordnete Bilten-Schänis Jud-Baumann Brigit, Schänis Paysen-Petersen Jacqueline, Rufi Baumgartner-Dürst Lukrezia, Bilten Kerenzen Schaub Walter, Obstalden Kamm-Menzi Marianne, Filzbach 78 Behörden der Kirchen Niederurnen Knöpfel Heini, Niederurnen Stuck Hans Markus, Niederurnen Fischli Elisabeth, Niederurnen Etter David, Niederurnen Hämmerli Christian, Niederurnen Mollis-Näfels Nöthiger-Lutz Markus, Mollis Schmid Brunner Ernst, Mollis Guler Verena, Mollis Perdrizat-Schweizer René, Mollis Kälin-Zimmermann Ruth, Mollis Senn Heidi, Mollis Netstal Cremonese-Feichtinger Andrea, Netstal Häuptli Saarah, Netstal -

35. Glarner Stadtlauf 2019, Glarus (173)
Datum: 01.11.19 35. Glarner Stadtlauf 2019, Glarus Zeit: 08:55:45 Seite: 1 (173) Schulklassen 3 Rang Teamname Klassierte Schnitt netto Diff zu 8.44,7 Name und Vorname Land/Ort Zeit Stnr 1. Engi Primarschule 5./6. Kl. 6 8:46.5 1.8 Bäbler Nevio Engi 7.55,2 1001 Frick Sven Elm 7.56,3 1003 Tschudi Sales Matt 8.16,2 1007 Blumer Anja Engi 8.45,0 1002 Lissner Emelie Engi 9.44,5 1004 Lüthi Flurina Engi 10.02,0 1005 2. Niederurnen Linth Escher Kl. 5c 13 8:38.8 5.9 Schmid Sheila Niederurnen 6.38,3 1451 Moos Noël Niederurnen 6.41,1 1445 Gübeli Lyra Niederurnen 7.04,2 1438 Menzi Matias Niederurnen 7.14,4 1444 Feusi Sonja Niederurnen 7.36,1 1436 Kovacevic Imran Niederurnen 8.22,4 1441 Maris Luisa-Maria Bilten 8.51,2 1443 Thoma Manuel Niederurnen 9.11,1 1452 Lazic Maria Ziegelbrücke 9.13,2 1442 Fontana Seraina Niederurnen 10.14,5 1437 Prenka Alexander Niederurnen 10.17,4 1447 Popadic Andjela Niederurnen 10.29,1 1446 Rrahmanaj Bleart Niederurnen 10.32,3 1448 3. Netstal Primarschule 5./6. Kl. 13 8:30.4 14.3 Correia Mariana Coutinho Netstal 7.01,6 1092 Fernandes Afonso Da Cunha Netstal 7.11,4 1093 Islami Venis Netstal 7.20,0 1094 Malacarne Moira Netstal 7.38,8 1098 Stombellini Lorenzo Netstal 7.55,5 1102 Sahinovic Aldin Netstal 8.08,8 1101 Ajeti Endrita Netstal 8.46,1 1089 Rodrigues Kevin Netstal 8.48,3 1100 Konuk Latife Netstal 9.15,4 1097 Kelmedi Blerijana Netstal 9.19,3 1096 Ramadani Diana Netstal 9.34,9 1099 Jenni Mark Netstal 9.47,7 1095 Barco Ruben Netstal 9.48,2 1091 4. -

Basisdokumentation Medien
Basisdokumentation Medien Blick von Ennetrösligen auf den Vorderglärnisch Bild: Kanton Glarus, Samuel Trümpy Photography In der dynamischen, weltoffenen und kulturell interessanten Gemeinde lässt es sich gut leben, arbeiten und geniessen. Nur eine Auto- und Bahn- stunde vor Zürichs Toren leben am Fusse einer imposanten Bergkulisse 12'461 Menschen (Stand 1. Januar 2020) in einer intakten Umwelt. Private und öffentliche Arbeitgeber bieten über 7’000 Arbeitsplätze an. Direkt um die Ecke sind Detaillisten, Cafés und Restaurants sowie Museen, Konzert- lokale und Bibliotheken. Eine Vielzahl von Vereinen, privaten Anbietern sowie öffentlichen Sport- und Freizeitanlagen prägen das reiche kulturelle Leben und Freizeitangebot. Kantonsschule und Kantonsspital befinden sich im Kantonshauptort. Naturräume wie das Klöntal, Aeugsten oder Schlatt erhält die Gemeinde bewusst. glarus.ch Medienkontakt Gemeinde Glarus – einzigartig vielseitig. Sonja Kohler Müller, Leiterin Standortförderung und Kommunikation Gemeindehausplatz 5, 8750 Glarus Telefon: +41 (0)58 611 81 27, E-Mail: [email protected] www.glarus.ch Pressebilder: www.flickr.com/glarus 02.03.2020 / Registratur: 2019-277 Inhaltsverzeichnis Facts & Figures 3 Politik 4 Organisation 7 Wirtschaft 8 Lebensraum 13 Familien 19 Bildung 20 Kultur & Gesellschaft 22 Freizeit & Tourismus 27 Ortsteile 31 2 Facts & Figures Fläche: 10'720 Hektaren davon Wald: 3'467 Hektaren Höchster Punkt: Bächistock, 2’914 m. ü. M Tiefster Punkt: Papierfabrik Netstal, 448 m. ü. M Einwohner: 12'461 (1.1.2020) Haushalte: 6’000 Arbeitsplätze: -

2020 11 24 Recycling Kalender 2021.Pdf
Fotos: Hans Bühler, Netstal Für Fragen und Auskünfte steht Ihnen die Abteilung Unterhaltsdienst (058 611 81 71) gerne zur Verfügung. Beachten Sie auch unsere Informationen im Internet auf www.gemeinde.glarus.ch. Im Onlineschalter: 'Entsorgung' finden Sie weitere Dokumente zu Gebühren und Verordnungen. Interessierte lernen an diesem Tag die Kunst des Kompostierens im eigenen Garten in Theorie und Praxis. Samstag 29. Mai 2021 (nähere Infos folgen) Die Gemeinde Glarus unterstützt die Aktion auch in diesem Jahr. Sie freut sich auf viele motivierte Teilnehmende, die ein Zeichen gegen Littering setzen und sich für eine saubere Gemeinde engagieren möchte. Bei der Interessengemeinschaft für eine saubere Umwelt (IGSU), welche den Clean-UP-Day in der Schweiz lanciert, können sich Vereine, Firmen, etc. direkt für die Teilnahme an dieser Sammelaktion anmelden. weitere Informationen unter: www.clean-up-day.ch Hauskehricht in zugebundenen, BRENNBARER gebührenpflichtigen, hellgrauen Kehrichtsäcken KEHRICHT 'Glarnerland' Bitte erst am Abfuhrtag bereitstellen! jeweils Dienstag + Freitag Sperrgut mit Sperrgutmarken versehen ab 7.00 Uhr SPERRGUT Ausmass max. 150 x 100 x 50 cm nur brennbares Material: Gewicht max. 15 kg (= 1 Marke) Loses Material binden und mit Marken versehen. Holz, Möbel, Bücher, An Feiertagen Es finden keine separaten Sperrgutabfuhren statt keine Abfuhr! Bauholz, Teppiche, Ski - nur saubere Zeitungen, Zeitschriften, Werbe- und Sammlungen durch Vereine Reklameprodukte siehe separate Daten auf - Papier mit Schnüren bündeln (nicht zu schwer) ALTPAPIER Rückseite Keine Papiersäcke, Karton, Plastik oder Gemeindesammelstellen Tetrapackungen An den Sammelstellen stehen Presscontainer für die Kartonsammlung bereit. Diese dürfen von KARTON Gemeindesammelstellen Privatpersonen und kleineren Betrieben benutzt werden. Nach Möglichkeit im eigenen Garten Mittwoch: kompostieren. Glarus + Netstal alle BIO - ABFÄLLE Alle pflanzlichen Garten- und Küchenabfälle im Grüngut aus Garten, grünen Container bereitstellen. -
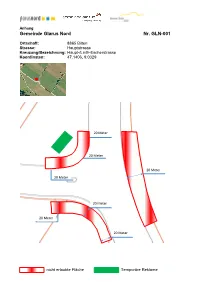
Gemeinde Glarus Nord Nr. GLN-001
Anhang Gemeinde Glarus Nord Nr. GLN-001 Ortschaft: 8865 Bilten Strasse: Hauptstrasse Kreuzung/Bezeichnung: Haupt-/Linth-Escherstrasse Koordinaten: 47.1406, 9.0329 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter nicht erlaubte Fläche Temporäre Reklame Gemeinde Glarus Nord Nr. GLN-002 Ortschaft: 8865 Bilten Strasse: Linth-Escherstrasse Kreuzung/Bezeichnung: Höhe SmartMarkt Koordinaten: 47.1493, 9.0323 5 Meter nicht erlaubte Fläche Temporäre Reklame Reg. Nr.: 33.03 / Seite 2/2 Gemeinde Glarus Nord Nr. GLN-003 Ortschaft: 8865 Bilten Strasse: Landstrasse Kreuzung/Bezeichnung: Land-/Niederrietstrasse Koordinaten: 47.1602, 9.0026 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter 20 Meter nicht erlaubte Fläche Temporäre Reklame Reg. Nr.: 33.03 / Seite 2/2 Gemeinde Glarus Nord Nr. GLN-004 Ortschaft: 8867 Niederurnen Strasse: Badstrasse Kreuzung/Bezeichnung: gegenüber Schiesstand Koordinaten: 47.1319, 9.0480 5 Meter nicht erlaubte Fläche Temporäre Reklame Reg. Nr.: 33.03 / Seite 2/2 Gemeinde Glarus Nord Nr. GLN-005 Ortschaft: 8867 Niederurnen Strasse: Ziegelbrückstrasse Kreuzung/Bezeichnung: Wiese römisch-katholische Kirche Koordinaten: 47.1276, 9.0554 3 Meter Aussenkante Trottoir nicht erlaubte Fläche Temporäre Reklame Reg. Nr.: 33.03 / Seite 2/2 Gemeinde Glarus Nord Nr. GLN-006 Ortschaft: 8867 Niederurnen Strasse: Hauptstrasse Kreuzung/Bezeichnung: Hauptstrasse / Gerbiweg Koordinaten: 47.1211, 9.0544 3 Meter Aussenkante Trottoir B a nicht erlaubte Fläche Temporäre Reklame Reg. Nr.: 33.03 / Seite 2/2 Gemeinde Glarus Nord Nr. GLN-007 Ortschaft: 8867 Niederurnen Strasse: Eternitstrasse Kreuzung/Bezeichnung: Haupt-/Eternitstrasse Koordinaten: 47.1199, 9.0581 5 Meter 20 Meter 10 Meter 20 Meter ab Aussenradius 20 Meter nicht erlaubte Fläche Temporäre Reklame Reg. -

Bahnhof Ennenda 13 Dezember 2020 - 11 Dezember 2021
Abfahrt Départ - Partenza - Departure Bahnhof Ennenda 13 Dezember 2020 - 11 Dezember 2021 4 00 13 00 22 00 4 47 S25 Linthal via Mitlödi–Schwanden GL– 13 09 S25 Zürich HB via Glarus–Netstal– 22 17 S6 Linthal via Mitlödi–Schwanden GL– Nidfurn-Haslen–Leuggelbach Näfels-Mollis–Nieder- und Oberurnen Nidfurn-Haslen–Leuggelbach 13 17 S6 Schwanden GL via Mitlödi 22 39 S6 Rapperswil SG via Glarus–Netstal– 5 00 13 39 S6 Rapperswil SG via Glarus–Netstal– Näfels-Mollis–Nieder- und Oberurnen Näfels-Mollis–Nieder- und Oberurnen 5 09 S25 Zürich HB via Glarus–Netstal– 13 47 S25 Linthal via Mitlödi–Schwanden GL– 23 00 Ziegelbrücke–Pfäffikon SZ Nidfurn-Haslen–Leuggelbach 5 17 S6 Schwanden GL via Mitlödi 23 17 S6 Linthal via Mitlödi–Schwanden GL– 5 39 S6 Rapperswil SG via Glarus–Netstal– 14 00 Nidfurn-Haslen–Leuggelbach Näfels-Mollis–Nieder- und Oberurnen 23 39 S6 Ziegelbrücke via Glarus–Netstal– 5 47 S25 Linthal via Mitlödi–Schwanden GL– 14 09 S25 Zürich HB via Glarus–Netstal– Näfels-Mollis–Nieder- und Oberurnen Nidfurn-Haslen–Leuggelbach Näfels-Mollis–Nieder- und Oberurnen 14 17 S6 Schwanden GL via Mitlödi 0 00 6 00 14 39 S6 Rapperswil SG via Glarus–Netstal– Näfels-Mollis–Nieder- und Oberurnen 0 17 S6 Schwanden GL via Mitlödi 6 09 S25 Zürich HB via Glarus–Netstal– 14 47 S25 Linthal via Mitlödi–Schwanden GL– 0 39 S6 Ziegelbrücke via Glarus–Netstal– Näfels-Mollis–Nieder- und Oberurnen Nidfurn-Haslen–Leuggelbach Näfels-Mollis–Nieder- und Oberurnen 6 17 S6 Schwanden GL via Mitlödi 6 39 S6 Rapperswil SG via Glarus–Netstal– 15 00 1 00 Näfels-Mollis–Nieder- und Oberurnen 6 47 S25 Linthal via Mitlödi–Schwanden GL– 15 09 S25 Zürich HB via Glarus–Netstal– 1 56 736 Linthal, Bahnhof via Nidfurn-Haslen–Leuggelbach Näfels-Mollis–Nieder- und Oberurnen Mitlödi, Hauptstrasse– 15 17 S6 Schwanden GL via Mitlödi Schwanden GL, Hauptstrasse– 7 00 15 39 S6 Rapperswil SG via Glarus–Netstal– Schwanden GL, Spittel– Näfels-Mollis–Nieder- und Oberurnen Nidfurn, Abzw. -

G Eschäftsbericht 20 14
Geschäftsbericht 2014 Inhaltsverzeichnis 3 Vorwort des Verwaltungsrats präsidenten 5 Vorwort des Geschäftsführers 7 Facts 9 Gesellschaftsorgane 10 Personelles 11 Jahresbericht Strom- und Kommunikations-Netz 13 Jahresbericht Erdgas 16 Jahresbericht Wasser 18 Jahresbericht Energie und Marketing 20 Neu : Naturstrom aus unserer Region 21 Luftseilbahn Luchsingen – Brunnenberg – Erholung vor der Haustür 23 Statistik 2014 25 Bilanz per 31. Dezember 2014 29 Erfolgsrechnung 2014 30 Bericht der Revisionsstelle 32 4 Reto Frey Verwaltungsratspräsident der tb.glarus Vorwort des Verwaltungsrats präsidenten Zukunft gestalten 5 Nach der Konsolidierungsphase als tb.glarus während Bereits während der Sommerferien 2014 wurde mit dem der ersten vier Jahre 2011– 2014 wurden insbesondere Bau der ersten Etappe von Wärmeverbund Glarus 1 be- im vergangenen Jahr massgebende Weichen für eine gonnen. Auch hier schreiten die Arbeiten termingerecht erfolgreiche Zukunft der tb.glarus in den kommenden voran und der VR darf mit grosser Genugtuung fest- Jahren gestellt. stellen, dass bereits 75 % der Versorgungsleistung des Wärmeverbunds aufgrund von Verträgen und Absichts- Nachdem im Bereich der Elektrizitätsversorgung die erklärungen vergeben sind. Netzkosten aufgrund der innerhalb eines halben Jahres erstellten Anlagenbuchhaltung reglementskonform be- Erfreulicherweise hat die Gemeindeversammlung bei rechnet werden konnten, wurde im Jahre 2014 auch die der Budgetberatung 2015 die Investitionen für die Er- Netzkostenberechnung im Bereich Erdgas-Versorgung schliessung von Glarus Süd mit Erdgas gutgeheissen. mittels des Systems « Nemo » eingeführt und zertifiziert. Zurzeit befinden wir uns noch in Verhandlungen mit der Dadurch sind nun auch im Bereich Gas die Grundlagen Gemeinde Glarus Süd und sind zuversichtlich, dass für die zukünftige Aufteilung der Preise in die Sparten dieses Projekt auch realisiert werden kann. Energie und Netzkosten erstellt. -

Erschliessung Bauzonen Impressum
Gemeinde Glarus Nord Revision Ortsplanung NUP II Entwurf Erschliessung Bauzonen Impressum Projekt Glarus Nord, Revision Ortsplanung NUP II Projektnummer: 27083 Dokument: Erschliessung Bauzonen Auftraggeber Gemeinde Glarus Nord Bearbeitungsstand Stand: Entwurf Bearbeitungsdatum: 11. Juni 2020, aktualisiert 24. Juni 2020 Druckdatum: 24. Juni 2020 Bearbeitung Schneiter Verkehrsplanung AG, Ennenda / Zürich (Marc Schneiter) STW AG für Raumplanung, Chur (Martin Reich) z:\gemeinde\glarus nord\29087_erschliessung_bauzonen_nup_ii\01_rap\02_resultate\04_pmb\20200624_erschliessung_bauzonen_1.docx Zusammenfassung In vorliegendem Bericht sind folgende Themen, als Grundlage und Nachweis zur Nutzungsplanung (NUP II) bearbeitet: - Es wird die heutige Erschliessungsfunktion aufgezeigt - Es werden Handlungsfelder identifiziert - Massnahmen zu künftigen Erschliessungen werden aufgearbei- tet - In Fokusthemen werden ausgewählte Fragestellungen vertieft Die Bearbeitung stützt sich auf sechs Leitsätze: 1. Es sind Verkehrsnetze für alle Verkehrsarten zu gewährleisten: Motorisierter Individualverkehr (MIV), öffentlicher Verkehr (ÖV), Fuss- und Veloverkehr 2. Das ÖV-Angebot wird sich in den nächsten Jahren verändern; es soll besser auf die Siedlungsentwicklung abgestimmt werden. 3. Strasseneigentum und tatsächliche Netzhierarchie zwischen Kanton und Gemeinde soll bereinigt werden. 4. Investitionen im untergeordneten Netz sind ressourcenbewusst vorzunehmen. 5. Eine massvolle Innenentwicklung soll auch in den gewachsenen Ortskernen ermöglicht werden. 6. Die Netze -

SWITZERLAND Andreas Tunger-Zanetti Introduction
SWITZERLAND Andreas Tunger-Zanetti1 Introduction “Swiss feel threatened by Islam” was the headline of the Sunday tabloid newspaper Sonntags-Blick on 27 August 2017.2 The claim was supported by a survey and can be seen as expressing in a nutshell a widespread perception in Switzerland. A much more nuanced picture was shown a few days earlier by the Religionsmonitor, a much more comprehensive survey by the German Bertelsmann Foundation, aptly condensed into the title “Integrated, but not accepted?”3 While the article under the first headline explicitly makes reference to unacceptable speeches in some mosques and to the radicalisation of young Muslims, the second expresses a broader range of issues. For Muslims and Islam in Switzerland, the year 2017 was marked by a constant oscillation between these two poles of a gloomy picture prevalent in media reporting, on the one hand, and an inconspicuous normality, on the other. The debate on “radicalisation”, supposed hate speech and Islamist activities, was stoked by a few new cases, such as that of a Libyan imam in Biel-Bienne. Regularly, proposals in the political debate asking, for example, for a stricter control of imams and the financing of mosques, are quick to follow, and with this the “burqa ban initiative” was successfully launched. The combination of a representative 1 Andreas Tunger-Zanetti is the coordinator of the Centre for Research on Religion at the University of Lucerne. 2 Faki, Sermîn, and Florian Blumer, “Schweizer fühlen sich vom Islam bedroht”, Sonntagsblick, 27 August 2017, pp. 2-3. 3 Halm, Dirk, and Martina Sauer, Muslime in Europa: integriert, aber nicht akzeptiert? (Gütersloh: Bertelsmann Stiftung, 2017), https://www.bertelsmann- stiftung.de/de/publikationen/publikation/did/muslime-in-europa/, accessed 22 April 2018. -

Pfarreiblatt Vom 11.05.-02.06.2019
Nr. 7 12./19./26. Mai 2019 1. Jahrgang Fridolin Druck und Medien AZA 8752 Näfels Herausgeber: Kath. Dekanat Glarus Redaktion: die jeweiligen Pfarrämter, Erscheint 17mal im Jahr. Adressänderungen an das GLARNER Wohnorts-Pfarramt melden! PFARREIBLATT für die katholischen Pfarreien Näfels, Oberurnen, Glarus, Netstal und für den Seelsorgeraum Glarus Süd Gedanken zum Hochfest CHRISTI HIMMELFAHRT NÄFELS Röm.-kath. Pfarramt St. Hilarius Denkmalweg 1 8752 Näfels Telefon 055 612 21 43 [email protected] www.kath-glarus.ch PC-Konto: 87-1946-9 Öffnungszeiten Montag 08.00 – 11.00 Uhr 15.00 – 18.00 Uhr Dienstag-Freitag 08.00 – 11.00 Uhr Seelsorger: Dekan Harald Eichhorn, Pfarrer [email protected] Dr. Sebastian Thayyil, Vikar Natel 078 735 81 12 [email protected] Rita Ming, Religionspädagogin Natel 079 344 51 02 17.25 Klosterkirche Rosenkranz 18.45 Pfarrkirche Beichtgelegenheit 18.45 Pfarrkirche Beichtgelegenheit bis 19.15 Br. René Fuchs OFM bis 19.15 19.30 Pfarrkirche Eucharistiefeier Seelsorger im Altersheim Letz 19.30 Pfarrkirche Eucharistiefeier Montag der 5. Osterwoche, 20. Mai Sekretariat: Montag, 13. Mai – Gedenktag Unserer 06.30 Klosterkirche Eucharistiefeier Wilma Kaspar-Hug Lieben Frau von Fatima 08.00 Pfarrkirche Eucharistiefeier 06.30 Klosterkirche Eucharistiefeier 15.00 Marienkirche Rosenkranz Religionsunterricht: 08.00 Pfarrkirche Eucharistiefeier Harald Eichhorn, Näfels 15.00 Marienkirche Rosenkranz Dienstag der 5. Osterwoche, 21. Mai Patricia Jacober, Oberurnen 06.30 Klosterkirche Eucharistiefeier Rita Ming, Näfels Dienstag der 4. Osterwoche, 14. Mai Edith Rast, Näfels 08.00 Pfarrkirche Eucharistiefeier 06.30 Klosterkirche Eucharistiefeier 09.30 Altersheim Letz Eucharistiefeier Kirchgemeindepräsidentin: 08.00 Pfarrkirche Eucharistiefeier 19.30 Klosterkirche Maiandacht Daniela Gallati-Landolt 09.30 Altersheim Letz Eucharistiefeier 19.30 Klosterkirche Maiandacht Mittwoch der 5. -

Bahn+Bus Glarnerland
SBB-RV-GL-A3-18.pdf 2 20.10.17 14:56 Wattwil | St.Gallen Meilen | Zürich S4 Uznach Rüti | Uster | Zürich Schmerikon S6 Kaltbrunn Kaltbrunn, Steinenbrücke Blumenau 521 Benken SG, 635 Rapperswil Post Nuolen 522 Benken SG Siebnen- 525 Wangen Tuggen, Post S4 S2 S2 S25 522 Schübelbach- Reichenburg S6 Lachen Buttikon 521 S27 Schänis, Rathausplatz Amden, Post Arvenbüel, Arven 524 S25 Schänis 635 650 Siebnen, Dorf Bilten, Ussbühl 523 Schulhaus Schübelbach, Buttikon SZ, Ziegelbrücke Pfäffikon SZ | Zürich Pfäffikon Bilten Reichenburg, Dorf Weesen, Post Oberstufenschule Bilten, S-Bahn Innerthal, Post/Gemeindezentrum Post Regionale Busse S-Bahn (nur zeitweise) Niederurnen, Ziegelbrückstr. Regionale Busse (nur zeitweise) Morgenholz Nieder- und Fernverkehr Niederurnen, Ochsenplatz Oberurnen S4 Mühlehorn Regionalverkehr Oberurnen, Feuerwehrplatz Andere Busse Näfels, Lintharena Seilbahn Mollis, Filzbach, Halt Fernverkehrszüge, S-Bahn und regionale Busse Kirchplatz511 Sportzentrum Sargans | Chur Halt S-Bahn und regionale Busse Näfels, Post Obstalden, Post Habergschwänd Halt nur in einer Richtung 518 Näfels-Mollis 512 Obersee Mullerenberg Näfels, Oberdorf Mollis, Bahnlinien Vorderdorf 501 502 519 S4 Uznach–Ziegelbrücke–Mühlehorn–Sargans–St.Gallen–Uznach S25 Fronalp (GL), Unter Stafel S6 Schwanden–Glarus–Ziegelbrücke–Uznach–Rapperswil Netstal, Wiggis Park S6 736 Linthal–Schwanden S6 Montag–Freitag als Bus 736 mit Anschluss an S6 in Schwanden Netstal, Altersheim Samstag/Sonntag teilweise als durchgehender Zug S6 (durchgehend Linthal–Rapperswil) Netstal S25 Linthal–Schwanden–Glarus–Ziegelbrücke–Zürich HB Glarus, Riedern GL, Kantonsschule/Spital S27 Ziegelbrücke–Siebnen-Wangen Staldengarten BRSB Linthal Braunwaldbahn–Braunwald (Standseilbahn) Klöntal, Riedern GL, Rhodannenberg 502 Schulhaus Glarus Glarus, Glarus, Pfrundhaus Rathaus Ennenda, Buslinien Klöntal,Klöntal, Richisau Vorauen Ennenda Gemeindehaus Aus Platzgründen sind im Plan nicht alle Bushaltestellen aufgeführt.