Species Belonging to Species
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Evolution of Genetic Systems in Filamentous Ascomycetes
Evolution of Genetic Systems in Filamentous Ascomycetes Evolutie van genetische systemen in hyphenvormende zakjeszwammen 0000 0513 3836 Promotor: dr. R.F. Hoekstra hoogleraar in de populatie- en kwantitatieve genetica fjtfoiißi f ßin Maarten J. Nauta Evolution of Genetic Systems in Filamentous Ascomycetes Proefschrift ter verkrijging van de graad van doctor in de landbouw- en milieuwetenschappen op gezag van de rector magnificus, dr. C.M. Karssen, in het openbaar te verdedigen op woensdag 12januar i 1994 des namiddags te vier uur in de Aula van de Landbouwuniversiteit te Wageningen. 15 0 S(p^ZJ> These investigations were supported by the Netherlands Organization for Scientific Research (N.W.O.). BibUt/FHEEK LAMDbOirWUNIVERSITEJi. WAGE NINGE N CIP-GEGEVENS KONINKLIJKE BIBLIOTHEEK, DEN HAAG Nauta, Maarten J. Evolution of genetic systems in filamentous ascomycetes / Maarten J. Nauta. - [ S.l. : s.n.]. -111 . Thesis Wageningen. - With ref. - With summary in Dutch. ISBN 90-5485-199-6 Subject headings: population genetics / ascomycetes. omslagontwerp: Ernst van Cleef foto omslag: Barrages tussen verschillende stammen van Podospora anserina als gevolg van vegetatieve incompatibiliteit. (met dank aan Inge Haspels) aan mijn ouders Voorwoord Dit proefschrift is het resultaat van vier jaar onderzoek, verricht bij de vakgroep Erfelijkheidsleer van de Landbouwuniversiteit in Wageningen. In zekere zin valt zo'n proefschrift te vergelijken met een levend wezen. Uit de genetica is bekend dat de verschijningsvorm van elk levend wezen tot stand komt door een combinatie van erfelijke aanleg en invloeden uit de omgeving. Voor een proefschrift geldt eigenlijk hetzelfde: Zowel het werk van de auteur, als de bijdragen van zijn omgeving zijn onontbeerlijk om tot een verschijningsvorm te komen. -

Taxonomy and Epidemiology of <I>Mucor Irregularis</I> , Agent Of
UvA-DARE (Digital Academic Repository) Taxonomy and epidemiology Mucor irregularis, agent of chronic cutaneous mucormycosis Lu, X.L.; Najafzadeh, M.J.; Dolatabadi, S.; Ran, Y.P.; Gerrits van den Ende, A.H.G.; Shen, Y.N.; Li, C.Y.; Xi, L.Y.; Hao, F.; Zhang, Q.Q.; Li, R.Y.; Hu, Z.M.; Lu, G.; Wang, J.J.; Drogari- Apiranthitou, M.; Klaassen, C.; Meis, J.F.; Hagen, F.; Liu, W.D.; de Hoog, G.S. DOI 10.3767/003158513X665539 Publication date 2013 Document Version Final published version Published in Persoonia - Molecular Phylogeny and Evolution of Fungi Link to publication Citation for published version (APA): Lu, X. L., Najafzadeh, M. J., Dolatabadi, S., Ran, Y. P., Gerrits van den Ende, A. H. G., Shen, Y. N., Li, C. Y., Xi, L. Y., Hao, F., Zhang, Q. Q., Li, R. Y., Hu, Z. M., Lu, G., Wang, J. J., Drogari-Apiranthitou, M., Klaassen, C., Meis, J. F., Hagen, F., Liu, W. D., & de Hoog, G. S. (2013). Taxonomy and epidemiology Mucor irregularis, agent of chronic cutaneous mucormycosis. Persoonia - Molecular Phylogeny and Evolution of Fungi, 30, 48-56. https://doi.org/10.3767/003158513X665539 General rights It is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons). Disclaimer/Complaints regulations If you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, stating your reasons. -

Algal Sex Determination and the Evolution of Anisogamy James Umen, Susana Coelho
Algal Sex Determination and the Evolution of Anisogamy James Umen, Susana Coelho To cite this version: James Umen, Susana Coelho. Algal Sex Determination and the Evolution of Anisogamy. Annual Review of Microbiology, Annual Reviews, 2019, 73 (1), 10.1146/annurev-micro-020518-120011. hal- 02187088 HAL Id: hal-02187088 https://hal.sorbonne-universite.fr/hal-02187088 Submitted on 17 Jul 2019 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Annu. Rev. Microbiol. 2019. 73:X–X https://doi.org/10.1146/annurev-micro-020518-120011 Copyright © 2019 by Annual Reviews. All rights reserved Umen • Coelho www.annualreviews.org • Algal Sexes and Mating Systems Algal Sex Determination and the Evolution of Anisogamy James Umen1 and Susana Coelho2 1Donald Danforth Plant Science Center, St. Louis, Missouri 63132, USA; email: [email protected] 2Sorbonne Université, UPMC Université Paris 06, CNRS, Algal Genetics Group, UMR 8227, Integrative Biology of Marine Models, Station Biologique de Roscoff, CS 90074, F-29688, Roscoff, France [**AU: Please write the entire affiliation in French or write it all in English, rather than a combination of English and French**] ; email: [email protected] Abstract Algae are photosynthetic eukaryotes whose taxonomic breadth covers a range of life histories, degrees of cellular and developmental complexity, and diverse patterns of sexual reproduction. -

Self-Fertility and Uni-Directional Mating-Type Switching in Ceratocystis Coerulescens, a Filamentous Ascomycete
Curr Genet (1997) 32: 52–59 © Springer-Verlag 1997 ORIGINAL PAPER T. C. Harrington · D. L. McNew Self-fertility and uni-directional mating-type switching in Ceratocystis coerulescens, a filamentous ascomycete Received: 6 July 1996 / 25 March 1997 Abstract Individual perithecia from selfings of most some filamentous ascomycetes. Although a switch in the Ceratocystis species produce both self-fertile and self- expression of mating-type is seen in these fungi, it is not sterile progeny, apparently due to uni-directional mating- clear if a physical movement of mating-type genes is in- type switching. In C. coerulescens, male-only mutants of volved. It is also not clear if the expressed mating-types otherwise hermaphroditic and self-fertile strains were self- of the respective self-fertile and self-sterile progeny are sterile and were used in crossings to demonstrate that this homologs of the mating-type genes in other strictly heter- species has two mating-types. Only MAT-2 strains are othallic species of ascomycetes. capable of selfing, and half of the progeny from a MAT-2 Sclerotinia trifoliorum and Chromocrea spinulosa show selfing are MAT-1. Male-only, MAT-2 mutants are self- a 1:1 segregation of self-fertile and self-sterile progeny in sterile and cross only with MAT-1 strains. Similarly, self- perithecia from selfings or crosses (Mathieson 1952; Uhm fertile strains generally cross with only MAT-1 strains. and Fujii 1983a, b). In tetrad analyses of selfings or crosses, MAT-1 strains only cross with MAT-2 strains and never self. half of the ascospores in an ascus are large and give rise to It is hypothesized that the switch in mating-type during self-fertile colonies, and the other ascospores are small and selfing is associated with a deletion of the MAT-2 gene. -

Sex in the Extremes: Lichen-Forming Fungi
Mycologist, Volume 19, Part 2 May 2005. ©Cambridge University Press Printed in the United Kingdom. DOI: 10.1017/S0269915XO5002016 Sex in the extremes: lichen-forming fungi FABIAN A. SEYMOUR, PETER D. CRITTENDEN & PAUL S. DYER* School of Biology, University of Nottingham, University Park, Nottingham, NG7 2RD, UK. Tel. +44 (0) 115 9513203, Fax +44 (0) 115 9513251 E-mail: [email protected]; [email protected] *Corresponding Author Lichens are characteristically found in environments subject to extremes of temperature, desiccation and low nutrient status. Despite this sexual structures are often formed in abundance. The underlying mechanisms of sex in lichen-forming fungi are discussed, together with possible ecological reasons for the persistence of sexuality. Special features of lichen sex are highlighted including sex at the limits of life on earth in Antarctica, re-licheniza- tion following sex and dispersal, and the perennial nature of lichen fruiting bodies. Keywords: lichen, fungi, sex, breeding system, (98%) belonging to the Ascomycotina (Kirk et al., symbiosis, extreme environments, Antarctica 2001). They display a variety of morphologies, from flattened crust (crustose) or leafy (foliose) forms to Lichens - living together in a long-term relation- shrubby or pendulous fruticose types (Honegger, 2001) ship (Figs 3, 4, 7, 8). Lichens are seen as a textbook example of a successful Life in extreme environments mutualistic symbiosis. They consist of at least two A key characteristic of lichens is that they have a organisms: a fungus (the ‘mycobiont’), and an remarkable ability to tolerate extreme environmental intimately associated photosynthetic partner (the conditions and sustain growth despite frequent cycles ‘photobiont’). -

<I>Mucorales</I>
Persoonia 30, 2013: 57–76 www.ingentaconnect.com/content/nhn/pimj RESEARCH ARTICLE http://dx.doi.org/10.3767/003158513X666259 The family structure of the Mucorales: a synoptic revision based on comprehensive multigene-genealogies K. Hoffmann1,2, J. Pawłowska3, G. Walther1,2,4, M. Wrzosek3, G.S. de Hoog4, G.L. Benny5*, P.M. Kirk6*, K. Voigt1,2* Key words Abstract The Mucorales (Mucoromycotina) are one of the most ancient groups of fungi comprising ubiquitous, mostly saprotrophic organisms. The first comprehensive molecular studies 11 yr ago revealed the traditional Mucorales classification scheme, mainly based on morphology, as highly artificial. Since then only single clades have been families investigated in detail but a robust classification of the higher levels based on DNA data has not been published phylogeny yet. Therefore we provide a classification based on a phylogenetic analysis of four molecular markers including the large and the small subunit of the ribosomal DNA, the partial actin gene and the partial gene for the translation elongation factor 1-alpha. The dataset comprises 201 isolates in 103 species and represents about one half of the currently accepted species in this order. Previous family concepts are reviewed and the family structure inferred from the multilocus phylogeny is introduced and discussed. Main differences between the current classification and preceding concepts affects the existing families Lichtheimiaceae and Cunninghamellaceae, as well as the genera Backusella and Lentamyces which recently obtained the status of families along with the Rhizopodaceae comprising Rhizopus, Sporodiniella and Syzygites. Compensatory base change analyses in the Lichtheimiaceae confirmed the lower level classification of Lichtheimia and Rhizomucor while genera such as Circinella or Syncephalastrum completely lacked compensatory base changes. -

Heterothallism and Potential Hybridization Events Inferred For
Du et al. IMA Fungus (2020) 11:4 https://doi.org/10.1186/s43008-020-0027-1 IMA Fungus RESEARCH Open Access Heterothallism and potential hybridization events inferred for twenty-two yellow morel species Xi-Hui Du1* , Dongmei Wu2, Heng Kang3*, Hanchen Wang1, Nan Xu1, Tingting Li1 and Keliang Chen1 Abstract Mating-type genes are central to sexual reproduction in ascomycete fungi and result in the establishment of reproductive barriers. Together with hybridization, they both play important roles in the evolution of fungi. Recently, potential hybridization events and MAT genes were separately found in the Elata Clade of Morchella. Herein, we characterized the MAT1–1-1 and MAT1–2-1 genes of twenty-two species in the Esculenta Clade, another main group in the genus Morchella, and proved heterothallism to be the predominant mating strategy among the twenty-two species tested. Ascospores of these species were multi-nuclear and had many mitochondrial nucleoids. The number of ascospore nuclei might be positively related with the species distribution range. Phylogenetic analyses of MAT1–1-1, MAT1–2-1, intergenic spacer (IGS), and partial histone acetyltransferase ELP3 (F1) were performed and compared with the species phylogeny framework derived from the ribosomal internal transcribed spacer region (ITS) and translation elongation factor 1-alpha (EF1-a) to evaluate their species delimitation ability and investigate potential hybridization events. Conflicting topologies among these genes genealogies and the species phylogeny were revealed and hybridization events were detected between several species. Different evolutionary patterns were suggested for MAT genes between the Esculenta and the Elata Clades. Complex evolutionary trajectories of MAT1–1-1, MAT1–2-1, F1 andIGSintheEsculentaCladewerehighlighted.Thesefindings contribute to a better understanding of the importance of hybridization and gene transfer in Morchella andespeciallyfortheappearanceofreproductivemodesduring its evolutionary process. -

Molecular Identification of Fungi
Molecular Identification of Fungi Youssuf Gherbawy l Kerstin Voigt Editors Molecular Identification of Fungi Editors Prof. Dr. Youssuf Gherbawy Dr. Kerstin Voigt South Valley University University of Jena Faculty of Science School of Biology and Pharmacy Department of Botany Institute of Microbiology 83523 Qena, Egypt Neugasse 25 [email protected] 07743 Jena, Germany [email protected] ISBN 978-3-642-05041-1 e-ISBN 978-3-642-05042-8 DOI 10.1007/978-3-642-05042-8 Springer Heidelberg Dordrecht London New York Library of Congress Control Number: 2009938949 # Springer-Verlag Berlin Heidelberg 2010 This work is subject to copyright. All rights are reserved, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilm or in any other way, and storage in data banks. Duplication of this publication or parts thereof is permitted only under the provisions of the German Copyright Law of September 9, 1965, in its current version, and permission for use must always be obtained from Springer. Violations are liable to prosecution under the German Copyright Law. The use of general descriptive names, registered names, trademarks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. Cover design: WMXDesign GmbH, Heidelberg, Germany, kindly supported by ‘leopardy.com’ Printed on acid-free paper Springer is part of Springer Science+Business Media (www.springer.com) Dedicated to Prof. Lajos Ferenczy (1930–2004) microbiologist, mycologist and member of the Hungarian Academy of Sciences, one of the most outstanding Hungarian biologists of the twentieth century Preface Fungi comprise a vast variety of microorganisms and are numerically among the most abundant eukaryotes on Earth’s biosphere. -
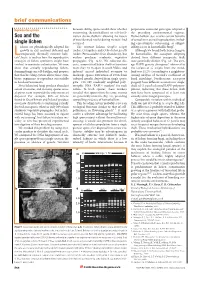
Brief Comms Layout 6/4Mx
brief communications Reproductive systems between sibling spores would show whether perpetuates successful genotypes adapted to outcrossing (heterothallism) or self-fertil- the prevailing environmental regimes. Sex and the ization (homothallism: allowing the fusion Homothallism also retains certain benefits of two identical nuclei during meiosis) had of sexual over asexual reproduction, includ- single lichen occurred. ing opportunistic outcrossing, as obligate ichens are physiologically adapted for The crustose lichens Graphis scripta selfing is rare in homothallic fungi8. growth in dry, nutrient-deficient and (order: Ostropales) and Ochrolechia parella Although we found both lichen fungi to Ltemporarily thermally extreme habi- (order: Pertusariales) fruit abundantly, but be homothallic, the ascospore offspring tats1, but it is unclear how the reproductive neither produces symbiotic vegetative derived from different conspecific thalli strategies of lichen symbionts might have propagules (Fig. 1a,b). We collected dis- were genetically distinct (Fig. 1d). The aver- evolved to maximize colonization. We now crete, symmetrical lichen thalli at locations age RAPD genetic divergence9 observed in show that sexually reproducing lichen- more than 10 m apart in south Wales, and five isolates of G. scripta from one wood- forming fungi can self-fertilize, and propose induced excised individual ascomata to land was 15.2% (according to a neighbour- that this breeding system allows these sym- discharge spores. Extraction of DNA from joining analysis of Jaccard’s coefficient of biotic organisms to reproduce successfully cultured mycelia derived from single spores band matching). Furthermore, ascospore in harsh environments. gave 218–263 randomly amplified poly- progeny from different ascomata on ‘single’ Most lichenized fungi produce abundant morphic DNA (RAPD) markers4 for each thalli of O. -

Discovery of a Sexual Cycle in Aspergillus Lentulus, a Close Relative of A
Discovery of a Sexual Cycle in Aspergillus lentulus, a Close Relative of A. fumigatus Sameira S. Swilaiman,a Céline M. O’Gorman,a S. Arunmozhi Balajee,b Paul S. Dyera School of Biology, University of Nottingham, University Park, Nottingham, United Kingdoma; Center for Global Health, Centers for Disease Control and Prevention, Atlanta, Georgia, USAb Aspergillus lentulus was described in 2005 as a new species within the A. fumigatus sensu lato complex. It is an opportunistic human pathogen causing invasive aspergillosis with high mortality rates, and it has been isolated from clinical and environmen- tal sources. The species is morphologically nearly identical to A. fumigatus sensu stricto, and this similarity has resulted in their frequent misidentification. Comparative studies show that A. lentulus has some distinguishing growth features and decreased in vitro susceptibility to several antifungal agents, including amphotericin B and caspofungin. Similar to the once-presumed-asex- ual A. fumigatus, it has only been known to reproduce mitotically. However, we now show that A. lentulus has a heterothallic sexual breeding system. A PCR-based mating-type diagnostic detected isolates of either the MAT1-1 or MAT1-2 genotype, and examination of 26 worldwide clinical and environmental isolates revealed similar ratios of the two mating types (38% versus 62%, respectively). MAT1-1 and MAT1-2 idiomorph regions were analyzed, revealing the presence of characteristic alpha and high-mobility-group (HMG) domain genes, together with other more unusual features such as a MAT1-2-4 gene. We then dem- onstrated that A. lentulus possesses a functional sexual cycle with mature cleistothecia, containing heat-resistant ascospores, being produced after 3 weeks of incubation. -

The Classification of Lower Organisms
The Classification of Lower Organisms Ernst Hkinrich Haickei, in 1874 From Rolschc (1906). By permission of Macrae Smith Company. C f3 The Classification of LOWER ORGANISMS By HERBERT FAULKNER COPELAND \ PACIFIC ^.,^,kfi^..^ BOOKS PALO ALTO, CALIFORNIA Copyright 1956 by Herbert F. Copeland Library of Congress Catalog Card Number 56-7944 Published by PACIFIC BOOKS Palo Alto, California Printed and bound in the United States of America CONTENTS Chapter Page I. Introduction 1 II. An Essay on Nomenclature 6 III. Kingdom Mychota 12 Phylum Archezoa 17 Class 1. Schizophyta 18 Order 1. Schizosporea 18 Order 2. Actinomycetalea 24 Order 3. Caulobacterialea 25 Class 2. Myxoschizomycetes 27 Order 1. Myxobactralea 27 Order 2. Spirochaetalea 28 Class 3. Archiplastidea 29 Order 1. Rhodobacteria 31 Order 2. Sphaerotilalea 33 Order 3. Coccogonea 33 Order 4. Gloiophycea 33 IV. Kingdom Protoctista 37 V. Phylum Rhodophyta 40 Class 1. Bangialea 41 Order Bangiacea 41 Class 2. Heterocarpea 44 Order 1. Cryptospermea 47 Order 2. Sphaerococcoidea 47 Order 3. Gelidialea 49 Order 4. Furccllariea 50 Order 5. Coeloblastea 51 Order 6. Floridea 51 VI. Phylum Phaeophyta 53 Class 1. Heterokonta 55 Order 1. Ochromonadalea 57 Order 2. Silicoflagellata 61 Order 3. Vaucheriacea 63 Order 4. Choanoflagellata 67 Order 5. Hyphochytrialea 69 Class 2. Bacillariacea 69 Order 1. Disciformia 73 Order 2. Diatomea 74 Class 3. Oomycetes 76 Order 1. Saprolegnina 77 Order 2. Peronosporina 80 Order 3. Lagenidialea 81 Class 4. Melanophycea 82 Order 1 . Phaeozoosporea 86 Order 2. Sphacelarialea 86 Order 3. Dictyotea 86 Order 4. Sporochnoidea 87 V ly Chapter Page Orders. Cutlerialea 88 Order 6. -

Topic: Heterothallism in Fungi
M.SC SEMESTER I : E CONTENT FOR MBOTCC-I: Phycology, Mycology and Bryology ,Unit 3 continued Faculty : Dr Tanuja, University Department of Botany, Patliputra University Topic: Heterothallism in fungi Ehrenbergh (1829), for the first time studied zygospores in the order Mucorales. The American mycologist Blakeslee (1904), reported that in the several genera of the order Mucorales, the zygospores are not formed at all. The term Heterothallism was first by A.F. Blakeslee in 1904 when he observed that zygospores could develop in some spp. only when two mycelia of different strains were allowed to come in contact with each other. According to Blakeslee (1904) Heterothallic condition is “essentially similar to that in dioecious plants and animals and although in this case the two complimentary individuals which are needed for sexual reproduction are in general not so conspicuously differentiated morphologically as in higher forms, such a morphological difference is often distinctly visible.” He also supported his view with facts and reasons, and also investigated that in the same order two types of species are found which may be named as homothallic and heterothallic species. When the two hyphae of the same mycelium produced by a single spore fuse with each other and a zygospore is developed, the species is said to be homothallic, e.g., Mucor hiemalis. Mucor mucedo and Mucor stolonifer are the typical heterothallic species. In heterothallic species the fusion can take place only among the different strained hyphae, which develop on different mycelia of different (+ and -) strains. In these species the zygospores cannot be produced by the fusion of two hyphae of the same strain.