RBI 6 English
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
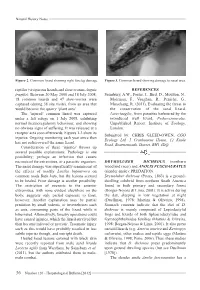
Reptiles (Viviparous Lizards and Slow-Worms Anguis REFERENCES Fragilis)
Natural History Notes Figure 2. Common lizard showing right foreleg damage. Figure 3. Common lizard showing damage to nasal area. reptiles (viviparous lizards and slow-worms Anguis REFERENCES fragilis). Between 20 May 2008 and 18 July 2008, Sainsbury, A.W., Foster, J., Bird, D., Moulton, N., 18 common lizards and 47 slow-worms were Molenaar, F., Vaughan, R., Peniche, G., captured (during 38 site visits), from an area that Marschang, R. (2011). Evaluating the threat to would become the quarry ‘plant area’. the conservation of the sand lizard, The ‘injured’ common lizard was captured Lacertaagilis, from parasites harboured by the under a felt refuge on 1 July 2008, exhibiting introduced wall lizard, Podarcismuralis. normal thermoregulatory behaviour, and showing Unpublished Report. Institute of Zoology, no obvious signs of suffering. It was released at a London. receptor area soon afterwards. Figures 1-3 show its Submitted by: CHRIS GLEED-OWEN, CGO injuries. Ongoing monitoring each year since then Ecology Ltd, 5 Cranbourne House, 12 Knole has not rediscovered the same lizard. Road, Bournemouth, Dorset, BH1 4DQ. Consideration of these ‘injuries’ throws up several possible explanations. Pathology is one possibility; perhaps an infection that causes necrosis of the extremities, or a parasitic organism. DRYMOLUBER DICHROUS (northern The nasal damage was superficially reminiscent of woodland racer) and ANOLIS FUSCOAURATUS the effects of toadfly Lucilia bufonivora on (slender anole): PREDATION. common toads Bufo bufo, but the lesions seemed Drymoluber dichrous (Peters, 1863) is a ground- to be healed. Frost damage is another possibility. dwelling colubrid from northern South America The restriction of necrosis to the anterior found in both primary and secondary forest extremities, with none evident elsewhere on the (Borges-Nojosa & Lima, 2001). -

Floristic Composition of Phytoplankton in Madeira River
G.J.B.A.H.S.,Vol.3(1):110-119 (January – March,2014) ISSN: 2319 – 5584 BIOLOGICAL DIVERSITY OF PHYTOPLANKTON AND PHYTOPERIPHYTON IN THE MADEIRA RIVER BASIN, AMAZON, BRAZIL Fabio Aprile1, Assad J. Darwich2, Pedro A. S. Mera2, Rosaluz Tavera3 & Karla K. do A. Serique2 1 Western of Pará Federal University - Ufopa. Av. Marechal Rondon s/n Santarém - PA 68040-070 Brazil. 2 National Institute of Research of Amazonian - Inpa. Av. André Araújo 2936 Manaus - AM 69060-001 Brazil. 3 Departamento de Ecología y Recursos Naturales, Universidad Nacional Autónoma de México, DF 04510 México. 4 Post-graduation Program of the National Institute of Research of Amazonian – Inpa. Abstract Floristic composition, abundance, richness and many other ecological patterns of phytoplankton and phytoperiphyton were studied in the Madeira River basin, between the Abunã River and Porto Velho City (Rondônia - Brazil), taking into account the hydrological period of the region. The results were correlated with abiotic factors, + + 2+ 2+ - 3- including the nutrients Na , K , Ca , Mg , N-NO3 , P-PO4 and chlorophyll-a. 283 taxa on the Madeira River and 327 taxa in the tributaries were identified belonging to six divisions and nine taxonomic classes. It was observed a considerable complexity in floristic composition, with a greater contribution from the division Chlorophyta, especially during the high water periods, when there was greater species richness recorded. Ecological indexes and statistical analysis confirmed strong seasonal pattern and differentiation between the species present in the white and clear waters. Keywords: floristic composition, diversity index, species richness, landscape ecology, Amazonia. 1. Introduction The Madeira River basin is located in the southwestern Amazonian region, and draining an enormous watershed frequently covered by Tropical Rain- Forest. -

A Comprehensive Multilocus Phylogeny of the Neotropical Cotingas
Molecular Phylogenetics and Evolution 81 (2014) 120–136 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev A comprehensive multilocus phylogeny of the Neotropical cotingas (Cotingidae, Aves) with a comparative evolutionary analysis of breeding system and plumage dimorphism and a revised phylogenetic classification ⇑ Jacob S. Berv 1, Richard O. Prum Department of Ecology and Evolutionary Biology and Peabody Museum of Natural History, Yale University, P.O. Box 208105, New Haven, CT 06520, USA article info abstract Article history: The Neotropical cotingas (Cotingidae: Aves) are a group of passerine birds that are characterized by Received 18 April 2014 extreme diversity in morphology, ecology, breeding system, and behavior. Here, we present a compre- Revised 24 July 2014 hensive phylogeny of the Neotropical cotingas based on six nuclear and mitochondrial loci (7500 bp) Accepted 6 September 2014 for a sample of 61 cotinga species in all 25 genera, and 22 species of suboscine outgroups. Our taxon sam- Available online 16 September 2014 ple more than doubles the number of cotinga species studied in previous analyses, and allows us to test the monophyly of the cotingas as well as their intrageneric relationships with high resolution. We ana- Keywords: lyze our genetic data using a Bayesian species tree method, and concatenated Bayesian and maximum Phylogenetics likelihood methods, and present a highly supported phylogenetic hypothesis. We confirm the monophyly Bayesian inference Species-tree of the cotingas, and present the first phylogenetic evidence for the relationships of Phibalura flavirostris as Sexual selection the sister group to Ampelion and Doliornis, and the paraphyly of Lipaugus with respect to Tijuca. -

Lições Das Interações Planta – Beija-Flor
UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE BIOLOGIA JÉFERSON BUGONI REDES PLANTA-POLINIZADOR NOS TRÓPICOS: LIÇÕES DAS INTERAÇÕES PLANTA – BEIJA-FLOR PLANT-POLLINATOR NETWORKS IN THE TROPICS: LESSONS FROM HUMMINGBIRD-PLANT INTERACTIONS CAMPINAS 2017 JÉFERSON BUGONI REDES PLANTA-POLINIZADOR NOS TRÓPICOS: LIÇÕES DAS INTERAÇÕES PLANTA – BEIJA-FLOR PLANT-POLLINATOR NETWORKS IN THE TROPICS: LESSONS FROM HUMMINGBIRD-PLANT INTERACTIONS Tese apresentada ao Instituto de Biologia da Universidade Estadual de Campinas como parte dos requisitos exigidos para a obtenção do Título de Doutor em Ecologia. Thesis presented to the Institute of Biology of the University of Campinas in partial fulfillment of the requirements for the degree of Doctor in Ecology. ESTE ARQUIVO DIGITAL CORRESPONDE À VERSÃO FINAL DA TESE DEFENDIDA PELO ALUNO JÉFERSON BUGONI E ORIENTADA PELA DRA. MARLIES SAZIMA. Orientadora: MARLIES SAZIMA Co-Orientador: BO DALSGAARD CAMPINAS 2017 Campinas, 17 de fevereiro de 2017. COMISSÃO EXAMINADORA Profa. Dra. Marlies Sazima Prof. Dr. Felipe Wanderley Amorim Prof. Dr. Thomas Michael Lewinsohn Profa. Dra. Marina Wolowski Torres Prof. Dr. Vinícius Lourenço Garcia de Brito Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no processo de vida acadêmica do aluno. DEDICATÓRIA À minha família por me ensinar o amor à natureza e a natureza do amor. Ao povo brasileiro por financiar meus estudos desde sempre, fomentando assim meus sonhos. EPÍGRAFE “Understanding patterns in terms of the processes that produce them is the essence of science […]” Levin, S.A. (1992). The problem of pattern and scale in ecology. Ecology 73:1943–1967. AGRADECIMENTOS Manifestar a gratidão às tantas pessoas que fizeram parte direta ou indiretamente do processo que culmina nesta tese não é tarefa trivial. -

Review of Fisheries Resource Use and Status in the Madeira River Basin (Brazil, Bolivia, and Peru) Before Hydroelectric Dam Completion
REVIEWS IN FISHERIES SCIENCE 2018, VOL. 0, NO. 0, 1–21 https://doi.org/10.1080/23308249.2018.1463511 Review of Fisheries Resource Use and Status in the Madeira River Basin (Brazil, Bolivia, and Peru) Before Hydroelectric Dam Completion Carolina R. C. Doria a,b, Fabrice Duponchelleb,c, Maria Alice L. Limaa, Aurea Garciab,d, Fernando M. Carvajal-Vallejosf, Claudia Coca Mendez e, Michael Fabiano Catarinog, Carlos Edwar de Carvalho Freitasc,g, Blanca Vegae, Guido Miranda-Chumaceroh, and Paul A. Van Dammee,i aLaboratory of Ichthyology and Fisheries – Department of Biology, Federal University of Rond^onia, Porto Velho, Brazil; bLaboratoire Mixte International – Evolution et Domestication de l’Ichtyofaune Amazonienne (LMI – EDIA), Iquitos (Peru), Santa Cruz (Bolivia); cInstitut de Recherche pour le Developpement (IRD), UMR BOREA (IRD-207, MNHN, CNRS-7208, Sorbonne Universite, Universite Caen Normandie, Universite des Antilles), Montpellier, France; dIIAP, AQUAREC, Iquitos, Peru; eFAUNAGUA, Institute for Applied Research on Aquatic Resources, Sacaba- Cochabamba, Plurinational State of Bolivia; fAlcide d’Orbigny Natural History Museum, ECOSINTEGRALES SRL (Ecological Studies for Integral Development and Nature Conservation), Cochabamba, Plurinational State of Bolivia; gDepartment of Fisheries Science, Federal University of Amazonas, Manaus, Brazil; hWildlife Conservation Society, Greater Madidi Tambopata Landscape Conservation Program, La Paz, Bolivia; iUnit of Limnology and Aquatic Resources (ULTRA), University of San Simon, Cochabamba, Bolivia -

Tinamiformes – Falconiformes
LIST OF THE 2,008 BIRD SPECIES (WITH SCIENTIFIC AND ENGLISH NAMES) KNOWN FROM THE A.O.U. CHECK-LIST AREA. Notes: "(A)" = accidental/casualin A.O.U. area; "(H)" -- recordedin A.O.U. area only from Hawaii; "(I)" = introducedinto A.O.U. area; "(N)" = has not bred in A.O.U. area but occursregularly as nonbreedingvisitor; "?" precedingname = extinct. TINAMIFORMES TINAMIDAE Tinamus major Great Tinamou. Nothocercusbonapartei Highland Tinamou. Crypturellus soui Little Tinamou. Crypturelluscinnamomeus Thicket Tinamou. Crypturellusboucardi Slaty-breastedTinamou. Crypturellus kerriae Choco Tinamou. GAVIIFORMES GAVIIDAE Gavia stellata Red-throated Loon. Gavia arctica Arctic Loon. Gavia pacifica Pacific Loon. Gavia immer Common Loon. Gavia adamsii Yellow-billed Loon. PODICIPEDIFORMES PODICIPEDIDAE Tachybaptusdominicus Least Grebe. Podilymbuspodiceps Pied-billed Grebe. ?Podilymbusgigas Atitlan Grebe. Podicepsauritus Horned Grebe. Podicepsgrisegena Red-neckedGrebe. Podicepsnigricollis Eared Grebe. Aechmophorusoccidentalis Western Grebe. Aechmophorusclarkii Clark's Grebe. PROCELLARIIFORMES DIOMEDEIDAE Thalassarchechlororhynchos Yellow-nosed Albatross. (A) Thalassarchecauta Shy Albatross.(A) Thalassarchemelanophris Black-browed Albatross. (A) Phoebetriapalpebrata Light-mantled Albatross. (A) Diomedea exulans WanderingAlbatross. (A) Phoebastriaimmutabilis Laysan Albatross. Phoebastrianigripes Black-lootedAlbatross. Phoebastriaalbatrus Short-tailedAlbatross. (N) PROCELLARIIDAE Fulmarus glacialis Northern Fulmar. Pterodroma neglecta KermadecPetrel. (A) Pterodroma -

Synopsis and Typification of Mexican and Central American
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Annalen des Naturhistorischen Museums in Wien Jahr/Year: 2018 Band/Volume: 120B Autor(en)/Author(s): Berger Andreas Artikel/Article: Synopsis and typification of Mexican and Central American Palicourea (Rubiaceae: Palicoureeae), part I: The entomophilous species 59-140 ©Naturhistorisches Museum Wien, download unter www.zobodat.at Ann. Naturhist. Mus. Wien, B 120 59–140 Wien, Jänner 2018 Synopsis and typification of Mexican and Central American Palicourea (Rubiaceae: Palicoureeae), part I: The entomophilous species A. Berger* Abstract The prominent but complex genus Psychotria (Rubiaceae: Psychotrieae) is one of the largest genera of flow- ering plants and its generic circumscription has been controversial for a long time. Recent DNA-phyloge- netic studies in combination with a re-evaluation of morphological characters have led to a disintegration process that peaked in the segregation of hundreds of species into various genera within the new sister tribe Palicoureeae. These studies have also shown that species of Psychotria subg. Heteropsychotria are nested within Palicourea, which was traditionally separated by showing an ornithophilous (vs. entomophilous) pol- lination syndrome. In order to render the genera Palicourea and Psychotria monophyletic groups, all species of subg. Heteropsychotria have to be transferred to Palicourea and various authors and publications have provided some of the necessary combinations. In the course of ongoing research on biotic interactions and chemodiversity of the latter genus, the need for a comprehensive and modern compilation of species of Pali courea in its new circumscription became apparent. As first step towards such a synopsis, the entomophilous Mexican and Central American species (the traditional concept of Psychotria subg. -

12. Plantas Y Líquenes Del Parque Nacional Natural De La Serranía De
PLANTAS Y LÍQUENES DEL PARQUE NACIONAL NATURAL SERRANÍA DE CHIRIBIQUETE, COLOMBIA Dairon Cárdenas1*1*,, MaríaMaría FernandaFernanda GonzálezGonzález12 ,, NóridaNórida MarínMarín11 y Sonia Sua11,, JulioJulio BetancurBetancur2 RESUMEN principales de formaciones vegetales registrados para la Guayana: bosques, matorrales, praderas o Se presenta el inventario de las plantas y los pastizales y la vegetación pionera sobre roca dura. líquenes del Parque Nacional Natural Serranía de En los afloramientos rocosos predominan especies Chiribiquete (PNNSCh), obtenido a través de como: Abolboda spp., Acanthella sprucei, Bonnetia información de herbarios y exploraciones de campo. sessilis, Calliandra vaupesiana, Clusia spp., Cyrilla Chiribiquete es el área protegida más grande de racemiflora, Decagonocarpus oppositifolius, Drosera Colombia, pertenece a la formación Guayana, esmeraldae, Euphronia hirtelloides, Gongylolepis tiene altitudes entre 200 y 800 metros y representa una gran meseta tectónica fraccionada por erosión martiana, Hevea nitida, Molongum lucidum, hídrica y disectada en numerosas mesas (tepuyes). Navia spp., Ochthocosmus berryi, Monotrema aemu- Se registraron 2,138 especies que corresponden lans, Pachira coriacea, Parahancornia surrogata, al 7.6 % de las encontradas en Colombia y al 66.5 Senefelderopsis chiribiquetensis, Steyerbromelia % de las de la región amazónica. Las familias con garcia-barrigae, Styrax rigidifolius, Tepuianthus más especies fueron: Fabaceae (184), Rubiaceae savannensis, Utricularia spp., Vellozia tubiflora (167) -

Reproductive Biology and Nest-Site Selection of the Mato Grosso Antbird Cercomacra Melanaria in the Brazilian Pantanal
Revista Brasileira de Ornitologia, 22(3), 270-277 ARTICLE September 2014 Reproductive biology and nest-site selection of the Mato Grosso Antbird Cercomacra melanaria in the Brazilian Pantanal Bianca Bernardon1, 2, 3, Paula Fernanda Albonette de Nóbrega1 and João Batista de Pinho1 1 Laboratório de Ornitologia, Núcleo de Estudos Ecológicos do Pantanal, Instituto de Biociências, Universidade Federal de Mato Grosso – UFMT, Avenida Fernando Corrêa, s/nº, Coxipó, CEP 78060-900, Cuiabá, MT, Brazil 2 Current Address: Grupo de Pesquisa em Ecologia de Vertebrados Terrestres, Instituto de Desenvolvimento Sustentável Mamirauá – IDSM, Estrada do Bexiga, 2584, CEP 69553-225, Tefé, AM, Brazil 3 Corresponding author: [email protected] Received on 27 December 2013. Accepted on 26 June 2014. ABSTRACT: Economic activities, such as extensive cattle ranching, may seriously threaten the types of forest most important to the Mato Grosso Antbird Cercomacra melanaria, and information on reproductive biology is essential for defining sound conservation and management strategies for the species. Here, we report on the reproductive biology of this species in the Brazilian Pantanal, focusing on attributes such as incubation and nestling periods, as well as characteristics of parental care. The hypothesis that nest occurrence is associated with canopy opening was also tested. Average nest height was 0.98 m; mean nest construction and egg incubation periods were 12.2 and 14 days, respectively; average nestling period was 9.4 days. Both males and females participated in nest construction, egg incubation, and feeding of nestlings. The canopy was consistently more open away from the nests (18.74%) than at nest sites (10.10%; P < 0.0001), indicating that C. -

Proyecto Mesozonificación Ecológica Y Económica Para
Mesozonificación Ecológica y Económica para el Desarrollo Sostenible de la Provincia de Satipo Informe temático Vegetación Filomeno Encarnación Cajañaupa / Ricardo Zárate Gómez Mesozonificación Ecológica y Económica para el Desarrollo Sostenible de la provincia de Satipo Informe temático: VEGETACIÓN / Filomeno Encarnación Cajañaupa Ricardo Zárate Gómez © Instituto de Investigaciones de la Amazonía Peruana Programa de Investigación en Cambio Climático, Desarrollo Territorial y Ambiente Av. José Abelardo Quiñones km 2,5 Teléfonos: (+51) (65) 265515 / 265516 Fax: (+51) (65) 265527 www.iiap.org.pe / [email protected] Iquitos-Perú, 2011 El presente estudio fue financiado con fondos del Plan de Impacto Rápido de Lucha contra las Drogas – PIR, canalizados por DEVIDA Cita sugerida: Encarnación, F. y Zárate, R. 2011. Vegetación, informe temático. Proyecto Mesozonificación Ecológica y Económica para el Desarrollo Sostenible de la provincia de Satipo, convenio entre el IIAP, DEVIDA y la Municipalidad Provincial de Satipo. Iquitos – Perú. La información contenida en este informe puede ser reproducida total o parcialmente siempre y cuando se mencione la fuente de origen. [MESOZONIFICACIÓN ECOLÓGICA Y ECONÓMICA PARA EL DESARROLLO SOSTENIBLE DE LA PROVINCIA DE SATIPO] VEGETACIÓN Contenido PRESENTACIÓN ................................................................................................ 5 RESUMEN ........................................................................................................ 6 I. OBJETIVOS ............................................................................................... -

Literature Cited in Lizards Natural History Database
Literature Cited in Lizards Natural History database Abdala, C. S., A. S. Quinteros, and R. E. Espinoza. 2008. Two new species of Liolaemus (Iguania: Liolaemidae) from the puna of northwestern Argentina. Herpetologica 64:458-471. Abdala, C. S., D. Baldo, R. A. Juárez, and R. E. Espinoza. 2016. The first parthenogenetic pleurodont Iguanian: a new all-female Liolaemus (Squamata: Liolaemidae) from western Argentina. Copeia 104:487-497. Abdala, C. S., J. C. Acosta, M. R. Cabrera, H. J. Villaviciencio, and J. Marinero. 2009. A new Andean Liolaemus of the L. montanus series (Squamata: Iguania: Liolaemidae) from western Argentina. South American Journal of Herpetology 4:91-102. Abdala, C. S., J. L. Acosta, J. C. Acosta, B. B. Alvarez, F. Arias, L. J. Avila, . S. M. Zalba. 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetologia 26 (Suppl. 1):215-248. Abell, A. J. 1999. Male-female spacing patterns in the lizard, Sceloporus virgatus. Amphibia-Reptilia 20:185-194. Abts, M. L. 1987. Environment and variation in life history traits of the Chuckwalla, Sauromalus obesus. Ecological Monographs 57:215-232. Achaval, F., and A. Olmos. 2003. Anfibios y reptiles del Uruguay. Montevideo, Uruguay: Facultad de Ciencias. Achaval, F., and A. Olmos. 2007. Anfibio y reptiles del Uruguay, 3rd edn. Montevideo, Uruguay: Serie Fauna 1. Ackermann, T. 2006. Schreibers Glatkopfleguan Leiocephalus schreibersii. Munich, Germany: Natur und Tier. Ackley, J. W., P. J. Muelleman, R. E. Carter, R. W. Henderson, and R. Powell. 2009. A rapid assessment of herpetofaunal diversity in variously altered habitats on Dominica. -

Southern Wing-Banded Antbird, Myrmornis Torquata Myrmornithinae
Thamnophilidae: Antbirds, Species Tree I Northern Wing-banded Antbird, Myrmornis stictoptera ⋆Southern Wing-banded Antbird, Myrmornis torquata ⋆ Myrmornithinae Spot-winged Antshrike, Pygiptila stellaris Russet Antshrike, Thamnistes anabatinus Rufescent Antshrike, Thamnistes rufescens Guianan Rufous-rumped Antwren, Euchrepomis guianensus ⋆Western Rufous-rumped Antwren, Euchrepomis callinota Euchrepomidinae Yellow-rumped Antwren, Euchrepomis sharpei Ash-winged Antwren, Euchrepomis spodioptila Chestnut-shouldered Antwren, Euchrepomis humeralis ⋆Stripe-backed Antbird, Myrmorchilus strigilatus ⋆Dot-winged Antwren, Microrhopias quixensis ⋆Yapacana Antbird, Aprositornis disjuncta ⋆Black-throated Antbird, Myrmophylax atrothorax ⋆Gray-bellied Antbird, Ammonastes pelzelni MICRORHOPIINI ⋆Recurve-billed Bushbird, Neoctantes alixii ⋆Black Bushbird, Neoctantes niger Rondonia Bushbird, Neoctantes atrogularis Checker-throated Stipplethroat, Epinecrophylla fulviventris Western Ornate Stipplethroat, Epinecrophylla ornata Eastern Ornate Stipplethroat, Epinecrophylla hoffmannsi Rufous-tailed Stipplethroat, Epinecrophylla erythrura White-eyed Stipplethroat, Epinecrophylla leucophthalma Brown-bellied Stipplethroat, Epinecrophylla gutturalis Foothill Stipplethroat, Epinecrophylla spodionota Madeira Stipplethroat, Epinecrophylla amazonica Roosevelt Stipplethroat, Epinecrophylla dentei Negro Stipplethroat, Epinecrophylla pyrrhonota Brown-backed Stipplethroat, Epinecrophylla fjeldsaai ⋆Napo Stipplethroat, Epinecrophylla haematonota ⋆Streak-capped Antwren, Terenura