Molecular Basis of Olfactory Chemoreception in the Common
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Medical Review Officer Manual
Department of Health and Human Services Substance Abuse and Mental Health Services Administration Center for Substance Abuse Prevention Medical Review Officer Manual for Federal Agency Workplace Drug Testing Programs EFFECTIVE OCTOBER 1, 2010 Note: This manual applies to Federal agency drug testing programs that come under Executive Order 12564 dated September 15, 1986, section 503 of Public Law 100-71, 5 U.S.C. section 7301 note dated July 11, 1987, and the Department of Health and Human Services Mandatory Guidelines for Federal Workplace Drug Testing Programs (73 FR 71858) dated November 25, 2008 (effective October 1, 2010). This manual does not apply to specimens submitted for testing under U.S. Department of Transportation (DOT) Procedures for Transportation Workplace Drug and Alcohol Testing Programs (49 CFR Part 40). The current version of this manual and other information including MRO Case Studies are available on the Drug Testing page under Medical Review Officer (MRO) Resources on the SAMHSA website: http://www.workplace.samhsa.gov Previous Versions of this Manual are Obsolete 3 Table of Contents Chapter 1. The Medical Review Officer (MRO)........................................................................... 6 Chapter 2. The Federal Drug Testing Custody and Control Form ................................................ 7 Chapter 3. Urine Drug Testing ...................................................................................................... 9 A. Federal Workplace Drug Testing Overview.................................................................. -

Chemoreception
Senses 5 SENSES live version • discussion • edit lesson • comment • report an error enses are the physiological methods of perception. The senses and their operation, classification, Sand theory are overlapping topics studied by a variety of fields. Sense is a faculty by which outside stimuli are perceived. We experience reality through our senses. A sense is a faculty by which outside stimuli are perceived. Many neurologists disagree about how many senses there actually are due to a broad interpretation of the definition of a sense. Our senses are split into two different groups. Our Exteroceptors detect stimulation from the outsides of our body. For example smell,taste,and equilibrium. The Interoceptors receive stimulation from the inside of our bodies. For instance, blood pressure dropping, changes in the gluclose and Ph levels. Children are generally taught that there are five senses (sight, hearing, touch, smell, taste). However, it is generally agreed that there are at least seven different senses in humans, and a minimum of two more observed in other organisms. Sense can also differ from one person to the next. Take taste for an example, what may taste great to me will taste awful to someone else. This all has to do with how our brains interpret the stimuli that is given. Chemoreception The senses of Gustation (taste) and Olfaction (smell) fall under the category of Chemoreception. Specialized cells act as receptors for certain chemical compounds. As these compounds react with the receptors, an impulse is sent to the brain and is registered as a certain taste or smell. Gustation and Olfaction are chemical senses because the receptors they contain are sensitive to the molecules in the food we eat, along with the air we breath. -

Understanding Sensory Processing: Looking at Children's Behavior Through the Lens of Sensory Processing
Understanding Sensory Processing: Looking at Children’s Behavior Through the Lens of Sensory Processing Communities of Practice in Autism September 24, 2009 Charlottesville, VA Dianne Koontz Lowman, Ed.D. Early Childhood Coordinator Region 5 T/TAC James Madison University MSC 9002 Harrisonburg, VA 22807 [email protected] ______________________________________________________________________________ Dianne Koontz Lowman/[email protected]/2008 Page 1 Looking at Children’s Behavior Through the Lens of Sensory Processing Do you know a child like this? Travis is constantly moving, pushing, or chewing on things. The collar of his shirt and coat are always wet from chewing. When talking to people, he tends to push up against you. Or do you know another child? Sierra does not like to be hugged or kissed by anyone. She gets upset with other children bump up against her. She doesn’t like socks with a heel or toe seam or any tags on clothes. Why is Travis always chewing? Why doesn’t Sierra liked to be touched? Why do children react differently to things around them? These children have different ways of reacting to the things around them, to sensations. Over the years, different terms (such as sensory integration) have been used to describe how children deal with the information they receive through their senses. Currently, the term being used to describe children who have difficulty dealing with input from their senses is sensory processing disorder. _____________________________________________________________________ Sensory Processing Disorder -

UTAH PESTS Staff
UTAH PESTS News Utah Plant Pest Diagnostic Laboratory and USU Extension Vol. IV, Winter 2010 Battling Bed Bugs in Utah “Sleep tight, don’t let the bed bugs bite.” All people know this phrase, and the harsh reality of its meaning is becom- What’s Inside ing known once again. Over the past Turfgrass Insect Pests of decade, reports of bed bugs (Cimicidae: Utah Cimex lectularius) throughout North America and abroad have been on the Encouraging Native Pol- linators in Your Yard and rise. Accordingly, bed bug submissions Garden to the UPPDL have also been increasing. This article will briefly explain the recent In the Spotlight: Are resurgence of bed bugs, and consider- Native Plants Resistant to ations for selecting a pest control com- Pests? bugwood.org pany to eradicate bed bug problems. On the Lookout for Invasive Tree Fruit and HISTORY OF BED BUGS Landscape Pests In the 1920s and 1930s, Americans were News, Publications, Web plagued by bed bugs. Some reports sites, Calendar stated that one out of every three homes was infested. People could pick News Highlights up unwanted bugs on buses, taxis, in the NEW UTAH PESTS movie theater, and just about anywhere. FACT SHEETS But in the early 1950s, bed bugs disap- bugwood.org The following can be peared from the developed world’s radar, found on our Web site: thanks to new insecticides like DDT, and Raspberry Horntail improved living standards. DDT applica- Community tions in homes, hotels, transportation Grasshopper Control vehicles, and health care facilities would kill bed bugs for several months to over a year. -

Functional Properties of Insect Olfactory Receptors: Ionotropic Receptors and Odorant Receptors
Cell and Tissue Research (2021) 383:7–19 https://doi.org/10.1007/s00441-020-03363-x REVIEW Functional properties of insect olfactory receptors: ionotropic receptors and odorant receptors Dieter Wicher1 · Fabio Miazzi2 Received: 14 August 2020 / Accepted: 19 November 2020 / Published online: 27 January 2021 © The Author(s) 2021 Abstract The majority of insect olfactory receptors belong to two distinct protein families, the ionotropic receptors (IRs), which are related to the ionotropic glutamate receptor family, and the odorant receptors (ORs), which evolved from the gustatory recep- tor family. Both receptor types assemble to heteromeric ligand-gated cation channels composed of odor-specifc receptor proteins and co-receptor proteins. We here present in short the current view on evolution, function, and regulation of IRs and ORs. Special attention is given on how their functional properties can meet the environmental and ecological challenges an insect has to face. Keywords Insect olfaction · Ionotropic receptor · Odorant receptor · Ion channel · Olfactory sensory neuron · Signal transduction · Sensitization · Adaptation Introduction receptors (IRs). The frst members of the OR family were discovered two decades ago (Clyne et al. 1999; Gao and The olfactory system is dedicated to detect and to encode Chess 1999; Vosshall et al. 1999), whereas the IRs that are information from volatile chemical signals. Such signals can related to ionotropic glutamate receptors were frst reported be categorized according to the information they transfer. ten years later (Benton et al. 2009). For example, chemosignals involved in social communica- While ORs solely detect volatile chemosignals, IRs tion may be informative solely for the receiver as an olfac- are multimodal receptive entities (Fig. -

The Olfactory Bulb Theta Rhythm Follows All Frequencies of Diaphragmatic Respiration in the Freely Behaving Rat
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Frontiers - Publisher Connector ORIGINAL RESEARCH ARTICLE published: 11 June 2014 BEHAVIORAL NEUROSCIENCE doi: 10.3389/fnbeh.2014.00214 The olfactory bulb theta rhythm follows all frequencies of diaphragmatic respiration in the freely behaving rat Daniel Rojas-Líbano 1,2†, Donald E. Frederick 2,3, José I. Egaña 4 and Leslie M. Kay 1,2,3* 1 Committee on Neurobiology, The University of Chicago, Chicago, IL, USA 2 Institute for Mind and Biology, The University of Chicago, Chicago, IL, USA 3 Department of Psychology, The University of Chicago, Chicago, IL, USA 4 Departamento de Anestesiología y Reanimación, Facultad de Medicina, Universidad de Chile, Santiago, Chile Edited by: Sensory-motor relationships are part of the normal operation of sensory systems. Sensing Donald A. Wilson, New York occurs in the context of active sensor movement, which in turn influences sensory University School of Medicine, USA processing. We address such a process in the rat olfactory system. Through recordings of Reviewed by: the diaphragm electromyogram (EMG), we monitored the motor output of the respiratory Thomas A. Cleland, Cornell University, USA circuit involved in sniffing behavior, simultaneously with the local field potential (LFP) of Emmanuelle Courtiol, New York the olfactory bulb (OB) in rats moving freely in a familiar environment, where they display University Langone Medical Center, a wide range of respiratory frequencies. We show that the OB LFP represents the sniff USA cycle with high reliability at every sniff frequency and can therefore be used to study the *Correspondence: neural representation of motor drive in a sensory cortex. -

Color, Taste, and Odor: What You Should Know
Color, Taste, and Odor: What you should know From time to time the MassDEP receives consumer questions or complaints regarding the look, taste or the odor of drinking water. Listed below are common problems with drinking water and their most common causes. Please note that a particular problem in your drinking water may be the result of a cause not listed here; the only way to confirm a cause is to have a certified lab analyze the water and discuss the results with drinking water professional. If you receive water from a public drinking water system it is important to contact the Public Water Supply (PWS) before having a laboratory analyze the water. Information on private water testing is available. Filtering or treating the water may remedy persistent problems; however MassDEP does not recommend filtering or treating your water supply if your water is supplied by a MassDEP- approved PWS. MassDEP also does not regulate or recommend specific treatment systems for private home use. If you decide to use a filtration or treatment device in your home, the Department strongly encourages you to contact National Sanitation Foundation (NSF) for a list of approved devices. If you purchase a treatment device for private home use MassDEP also strongly recommends that it is maintained and provide active maintenance according to the manufacturer's instructions. Failure to maintain the equipment properly may make treatment ineffective and/or may create the potential for contamination. Common problems with drinking water are grouped into three categories: Color problems Taste / odor problems Particles in water If the problem with your water is not described here, if you are on a public water system please contact the public water department in your city or town or the MassDEP Drinking Water Program at your nearest regional MassDEP office. -

Special Issue “Olfaction: from Genes to Behavior”
G C A T T A C G G C A T genes Editorial Special Issue “Olfaction: From Genes to Behavior” Edgar Soria-Gómez 1,2,3 1 Department of Neurosciences, University of the Basque Country UPV/EHU, 48940 Leioa, Spain; [email protected] or [email protected] 2 Achucarro Basque Center for Neuroscience, Science Park of the UPV/EHU, 48940 Leioa, Spain 3 IKERBASQUE, Basque Foundation for Science, 48013 Bilbao, Spain Received: 12 June 2020; Accepted: 15 June 2020; Published: 15 June 2020 The senses dictate how the brain represents the environment, and this representation is the basis of how we act in the world. Among the five senses, olfaction is maybe the most mysterious and underestimated one, probably because a large part of the olfactory information is processed at the unconscious level in humans [1–4]. However, it is undeniable the influence of olfaction in the control of behavior and cognitive processes. Indeed, many studies demonstrate a tight relationship between olfactory perception and behavior [5]. For example, olfactory cues are determinant for partner selection [6,7], parental care [8,9], and feeding behavior [10–13], and the sense of smell can even contribute to emotional responses, cognition and mood regulation [14,15]. Accordingly, it has been shown that a malfunctioning of the olfactory system could be causally associated with the occurrence of important diseases, such as neuropsychiatric depression or feeding-related disorders [16,17]. Thus, a clear identification of the biological mechanisms involved in olfaction is key in the understanding of animal behavior in physiological and pathological conditions. -

Altered Functional Properties of the Codling Moth Orco Mutagenized in the Intracellular Loop‑3 Yuriy V
www.nature.com/scientificreports OPEN Altered functional properties of the codling moth Orco mutagenized in the intracellular loop‑3 Yuriy V. Bobkov1, William B. Walker III2 & Alberto Maria Cattaneo1,2* Amino acid substitutions within the conserved polypeptide sequence of the insect olfactory receptor co‑receptor (Orco) have been demonstrated to infuence its pharmacological properties. By sequence analysis and phylogenetic investigation, in the Lepidopteran subgroup Ditrysia we identifed a fxed substitution in the intracellular loop‑3 (ICL‑3) of a conserved histidine to glutamine. By means of HEK293 cells as a heterologous system, we functionally expressed Orco from the Ditrysian model Cydia pomonella (CpomOrco) and compared its functional properties with a site‑directed mutagenized version where this ICL‑3‑glutamine was reverted to histidine (CpomOrcoQ417H). The mutagenized CpomOrcoQ417H displayed decreased responsiveness to VUAA1 and reduced response efcacy to an odorant agonist was observed, when co‑transfected with the respective OR subunit. Evidence of reduced responsiveness and sensitivity to ligands for the mutagenized Orco suggest the fxed glutamine substitution to be optimized for functionality of the cation channel within Ditrysia. In addition, contrary to the wild type, the mutagenized CpomOrcoQ417H preserved characteristics of VUAA‑binding when physiologic conditions turned to acidic. Taken together, our fndings provide further evidence of the importance of ICL‑3 in forming basic functional properties of insect Orco‑ and Orco/OR‑channels, and suggest involvement of ICL‑3 in the potential functional adaptation of Ditrysian Orcos to acidifed extra‑/intracellular environment. Te odorant receptor co-receptor, Orco, is a unique transmembrane protein, expressed in most of the olfac- tory sensory neurons (OSNs) of insect antennae1–3 and is highly conserved in sequence and function across all insects4,5. -
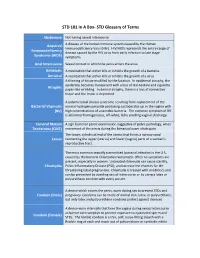
STD Glossary of Terms
STD 101 In A Box- STD Glossary of Terms Abstinence Not having sexual intercourse Acquired A disease of the human immune system caused by the Human Immunodeficiency Virus (HIV). HIV/AIDS represents the entire range of Immunodeficiency disease caused by the HIV virus from early infection to late stage Syndrome (AIDS) symptoms. Anal Intercourse Sexual contact in which the penis enters the anus. Antibiotic A medication that either kills or inhibits the growth of a bacteria. Antiviral A medication that either kills or inhibits the growth of a virus. A thinning of tissue modified by the location. In epidermal atrophy, the epidermis becomes transparent with a loss of skin texture and cigarette Atrophic paper-like wrinkling. In dermal atrophy, there is a loss of connective tissue and the lesion is depressed. A polymicrobial clinical syndrome resulting from replacement of the Bacterial Vaginosis normal hydrogen peroxide producing Lactobacillus sp. in the vagina with (BV) high concentrations of anaerobic bacteria. The common symptom of BV is abnormal homogeneous, off-white, fishy smelling vaginal discharge. Cervical Motion A sign found on pelvic examination suggestive of pelvic pathology; when Tenderness (CMT) movement of the cervix during the bimanual exam elicits pain. The lower, cylindrical end of the uterus that forms a narrow canal Cervix connecting the upper (uterus) and lower (vagina) parts of a woman's reproductive tract. The most common sexually transmitted bacterial infection in the U.S., caused by the bacteria Chlamydia trachomatis. Often no symptoms are present, especially in women. Untreated chlamydia can cause sterility, Chlamydia Pelvic Inflammatory Disease (PID), and increase the chances for life- threatening tubal pregnancies. -

Bacteria Associated with Larvae and Adults of the Asian Longhorned Beetle (Coleoptera: Cerambycidae)1
Bacteria Associated with Larvae and Adults of the Asian Longhorned Beetle (Coleoptera: Cerambycidae)1 John D. Podgwaite2, Vincent D' Amico3, Roger T. Zerillo, and Heidi Schoenfeldt USDA Forest Service, Northern Research Station, Hamden CT 06514 USA J. Entomol. Sci. 48(2): 128·138 (April2013) Abstract Bacteria representing several genera were isolated from integument and alimentary tracts of live Asian longhorned beetle, Anaplophora glabripennis (Motschulsky), larvae and adults. Insects examined were from infested tree branches collected from sites in New York and Illinois. Staphylococcus sciuri (Kloos) was the most common isolate associated with adults, from 13 of 19 examined, whereas members of the Enterobacteriaceae dominated the isolations from larvae. Leclercia adecarboxylata (Leclerc), a putative pathogen of Colorado potato beetle, Leptinotarsa decemlineata (Say), was found in 12 of 371arvae examined. Several opportunistic human pathogens, including S. xylosus (Schleifer and Kloos), S. intermedius (Hajek), S. hominis (Kloos and Schleifer), Pantoea agglomerans (Ewing and Fife), Serratia proteamaculans (Paine and Stansfield) and Klebsiella oxytoca (Fiugge) also were isolated from both larvae and adults. One isolate, found in 1 adult and several larvae, was identified as Tsukamurella inchonensis (Yassin) also an opportunistic human pathogen and possibly of Korean origin .. We have no evi dence that any of the microorganisms isolated are pathogenic for the Asian longhorned beetle. Key Words Asian longhorned beetle, Anaplophora glabripennis, bacteria The Asian longhorned beetle, Anoplophora glabripennis (Motschulsky) a pest native to China and Korea, often has been found associated with wood- packing ma terial arriving in ports of entry to the United States. The pest has many hardwood hosts, particularly maples (Acer spp.), and currently is established in isolated popula tions in at least 3 states- New York, NJ and Massachusetts (USDA-APHIS 201 0). -

A Pheromone Antagonist Liberates Female Sea Lamprey from a Sensory Trap to Enable Reliable Communication
A pheromone antagonist liberates female sea lamprey from a sensory trap to enable reliable communication Tyler J. Buchingera,1, Anne M. Scotta,1, Skye D. Fissettea, Cory O. Branta,2, Mar Huertasa,3,KeLia,4, Nicholas S. Johnsonb, and Weiming Lia,5 aDepartment of Fisheries and Wildlife, Michigan State University, East Lansing, MI 48824; and bHammond Bay Biological Station, US Geological Survey, Millersburg, MI 49759 Edited by John G. Hildebrand, University of Arizona, Tucson, AZ, and approved February 21, 2020 (received for review December 12, 2019) The evolution of male signals and female preferences remains a subsequent preference evolution precludes a full understanding central question in the study of animal communication. The sensory of how receiver biases shape communication. trap model suggests males evolve signals that mimic cues used Sea lamprey (Petromyzon marinus) are a useful model to track in nonsexual contexts and thus manipulate female behavior to the evolutionary trajectory of communication that originated via generate mating opportunities. Much evidence supports the sen- a sensory trap. Chemical cues and pheromones guide sea lam- sory trap model, but how females glean reliable information from prey behavior throughout their complex life history, especially both mimetic signals and their model cues remains unknown. We during the terminal reproductive phase (21). After parasitizing discovered a mechanism whereby a manipulative male signal guides fish in lakes or the Atlantic Ocean, prespawning sea lamprey reliable communication in sea lamprey (Petromyzon marinus). Mi- migrate into streams following chemical cues released by larvae gratory sea lamprey follow a larval cue into spawning streams; once residing in nursery habitats near spawning grounds.