Understanding the Structure of Flowers—The Wonderful Tool of Floral Formulae: a Response to Prenner & Al
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Only Write Down the Correct Answer Next to the Appropriate Question Number on Your Answer Sheet
BOT1B10– November 2015 FACULTY OF SCIENCE DEPARTMENT OF BOTANY and PLANT BIOTECHNOLOGY MODULE PLANT DIVERSITY BOT1B10 CAMPUS APK EXAMINATION NOVEMBER 2015 DATE SESSION 14/November/2015 08:30 – 11:30 EXAMINER: PROF A. MOTEETEE INTERNAL MODERATOR: MRS J. WILLIAMSON DURATION: 3 HOURS MARKS: 120 ____________________________________________________________________________________ NUMBER OF PAGES: 11 PAGES INSTRUCTIONS: ANSWER ALL THE QUESTIONS _____________________________________________________________________________ QUESTION 1 [10] Choose an answer that matches the question the best: Only write down the correct answer next to the appropriate question number on your answer sheet. 1.1. Apart from food and beverages, plants provide human beings with: a) Oxygen, nitrogen, construction materials b) Medicines, essential oils, oxygen, fuel c) Paper, wool, cotton, silk d) Herb and spices, carbon, sodium, fodder for animals 1 BOT1B10– November 2015 1.2 Gymnosperms bear their seeds on the surfaces of: a) Leaves b) Cones c) Stems d) Fruits 1.3 Bryophytes have life cycles that depend on what for reproduction? a) Water b) Soil c) Grass d) Sun 1.4 Plants that have xylem and phloem are known as: a) Seed plants b) Photosynthetic plants c) Vascular-plants d) Non-vascular plants 1.5 Which on the following is the stalk by which the leaf blade is attached to the stem? a) Peduncle b) Pedicel c) Inflorescence d) Petiole 1.6 Seed plants use __________ and ___________ to reproduce. a) Pollen and seed b) Seeds and water c) Food and water d) Leaves and petals 1.7 -
1Lecture Notes 2013
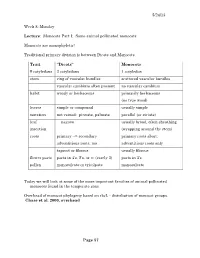
5/24/13 Week 8; Monday Lecture: Monocots Part I: Some animal pollinated monocots Monocots are monophyletic! Traditional primary division is between Dicots and Monocots Trait “Dicots” Monocots # cotyledons 2 cotyledons 1 cotyledon stem ring of vascular bundles scattered vascular bundles vascular cambium often present no vascular cambium habit woody or herbaceous primarily herbaceous (no true wood) leaves simple or compound usually simple venation net veined: pinnate, palmate parallel (or striate) leaf narrow usually broad, often sheathing insertion (wrapping around the stem) roots primary --> secondary primary roots abort; adventitious roots, too adventitious roots only taproot or fibrous usually fibrous flower parts parts in 4’s, 5’s, or ∞ (rarely 3) parts in 3’s pollen monosulcate or tricolpate monosulcate Today we will look at some of the more important families of animal pollinated monocots found in the temperate zone Overhead of monocot phylogeny based on rbcL - distribution of monocot groups. Chase et al. 2000, overhead Page 57 5/24/13 Lab only; limited discussion here. Show: “Plants are Cool, Too” video Araceae - Arum family (109 gen/2830 spp) 1) herbs (some epiphytes) 2) lvs simple or compound; broad and having an apparent petiole (‘pseudo-lamina’) development not same as in a dicot leaf blade 3) calcium oxalate crystals usually present – physical deterrent to herbivory 4) Inflorescence consisting of - spathe - bract (often colorful) surrounding the flowers - spadix - axis on which the flowers are borne (male above; female below, -

Plant Systematics Economic Botany and Ethnobotany
CORE PAPER- VIII PLANT SYSTEMATICS ECONOMIC BOTANY AND ETHNOBOTANY UNIT - III Rubiaceae Systematic position Class-Dicotyledons Sub class -Gamopetalae Series –Inferae Order - Rubiales Family-Rubiaceae Distribution of Rubiaceae: It is commonly known as Madder or Coffee family. It includes 6000 species and 500 genera. In India it is represented by 551 species. The members of this family are distributed in tropics, sub-tropics and temperate regions. Vegetative characters Habit and Habitatat. Trees -Adina cordifolia Shrubs- Gardenia (mostly), some are twinners- Paederia Climbers -Uncaria Herbs -Gallium Epiphytic eg Hymenopogon parasiticus Helophytic, or mesophytic, or xerophytic, or hydrophytic (Limnosipanea). Majority are perennials a few annuals, cultrivated as well as wild Root –branched tap root Stem- aerial,erect or weak, cylindrical or angular herbaceous Gallium or woody ,armed with spines Randia dementorum ,glabrous,pubescent hairy or smooth Stephegyne, branched, dichasial cymein Gallium. Leaf - Cauline and ramal Leaves stipulate. Stipules interpetiolar (between the petioles , or intrapetiolar; between the petiole and axis .leafy Gallium divided Borreria hair like Pentas sometimes fused to form a sheath GardeniaPetiolate, subsessile or sessile Gallium Leaves opposite Cinchona or whorled Gallium simple; Lamina entire; Cinchona opposite decussate Ixora ), reticulate Floral characters: Inflorescence- Flowers aggregated in ‘inflorescences’, or solitary (less often); in cymes, or in panicles, Cinchona or in heads (rarely, e.g. Morindeae, Gardenia). The ultimate inflorescence units compound cyme MussaendaInflorescences with involucral bracts (when capitate), or without involucral bracts; Flowers -Bracteate Gardenia ebracteate Cinchona Bracts persistant –Hymenopogan Pedicellate,subsessile Gardenia sessile RandinBracteolate or ebracteolate, complete or incomplete actinomorphic,, Rarely Zygomorphic Randeletin bisexual unisexual Coprosma , epigynous regular; mostly 4 merous, or 5 merous; cyclic; tetracyclic. -

Flowering Plant Families of Northwestern California: a Tabular Comparison
Humboldt State University Digital Commons @ Humboldt State University Botanical Studies Open Educational Resources and Data 12-2019 Flowering Plant Families of Northwestern California: A Tabular Comparison James P. Smith Jr Humboldt State University, [email protected] Follow this and additional works at: https://digitalcommons.humboldt.edu/botany_jps Part of the Botany Commons Recommended Citation Smith, James P. Jr, "Flowering Plant Families of Northwestern California: A Tabular Comparison" (2019). Botanical Studies. 95. https://digitalcommons.humboldt.edu/botany_jps/95 This Flora of Northwest California-Regional is brought to you for free and open access by the Open Educational Resources and Data at Digital Commons @ Humboldt State University. It has been accepted for inclusion in Botanical Studies by an authorized administrator of Digital Commons @ Humboldt State University. For more information, please contact [email protected]. FLOWERING PLANT FAMILIES OF NORTHWESTERN CALIFORNIA: A TABULAR COMPARISON James P. Smith, Jr. Professor Emeritus of Botany Department of Biological Sciences Humboldt State University December 2019 Scientific Name Habit Leaves Sexuality • Floral Formula Common Name Fruit Type • Comments Aceraceae TSV SC:O U-m [P] • K 4-5 C 4-5 A 4-10 G (2) Maple Paired samaras • leaves often palmately lobed Acoraceae H S:A U-m • P 3+3 A 6 or G (3) Sweet Flag Berry • aquatic; aromatic rhizomes Aizoaceae HS S:AO B • P [3] 5 [8] A 0-4 Gsi (2-5-4) Ice Plant Capsule (berry-like) • fleshy; stamens divided, petaloid Alismataceae -

NATIVE PLANT FIELD GUIDE Revised March 2012
NATIVE PLANT FIELD GUIDE Revised March 2012 Hansen's Northwest Native Plant Database www.nwplants.com Foreword Once upon a time, there was a very kind older gentleman who loved native plants. He lived in the Pacific northwest, so plants from this area were his focus. As a young lad, his grandfather showed him flowers and bushes and trees, the sweet taste of huckleberries and strawberries, the smell of Giant Sequoias, Incense Cedars, Junipers, pines and fir trees. He saw hummingbirds poking Honeysuckles and Columbines. He wandered the woods and discovered trillium. When he grew up, he still loved native plants--they were his passion. He built a garden of natives and then built a nursery so he could grow lots of plants and teach gardeners about them. He knew that alien plants and hybrids did not usually live peacefully with natives. In fact, most of them are fierce enemies, not well behaved, indeed, they crowd out and overtake natives. He wanted to share his information so he built a website. It had a front page, a page of plants on sale, and a page on how to plant natives. But he wanted more, lots more. So he asked for help. I volunteered and he began describing what he wanted his website to do, what it should look like, what it should say. He shared with me his dream of making his website so full of information, so inspiring, so educational that it would be the most important source of native plant lore on the internet, serving the entire world. -

Sinopsis De La Familia Acanthaceae En El Perú
Revista Forestal del Perú, 34 (1): 21 - 40, (2019) ISSN 0556-6592 (Versión impresa) / ISSN 2523-1855 (Versión electrónica) © Facultad de Ciencias Forestales, Universidad Nacional Agraria La Molina, Lima-Perú DOI: http://dx.doi.org/10.21704/rfp.v34i1.1282 Sinopsis de la familia Acanthaceae en el Perú A synopsis of the family Acanthaceae in Peru Rosa M. Villanueva-Espinoza1, * y Florangel M. Condo1 Recibido: 03 marzo 2019 | Aceptado: 28 abril 2019 | Publicado en línea: 30 junio 2019 Citación: Villanueva-Espinoza, RM; Condo, FM. 2019. Sinopsis de la familia Acanthaceae en el Perú. Revista Forestal del Perú 34(1): 21-40. DOI: http://dx.doi.org/10.21704/rfp.v34i1.1282 Resumen La familia Acanthaceae en el Perú solo ha sido revisada por Brako y Zarucchi en 1993, desde en- tonces, se ha generado nueva información sobre esta familia. El presente trabajo es una sinopsis de la familia Acanthaceae donde cuatro subfamilias (incluyendo Avicennioideae) y 38 géneros son reconocidos. El tratamiento de cada género incluye su distribución geográfica, número de especies, endemismo y carácteres diagnósticos. Un total de ocho nombres (Juruasia Lindau, Lo phostachys Pohl, Teliostachya Nees, Streblacanthus Kuntze, Blechum P. Browne, Habracanthus Nees, Cylindrosolenium Lindau, Hansteinia Oerst.) son subordinados como sinónimos y, tres especies endémicas son adicionadas para el país. Palabras clave: Acanthaceae, actualización, morfología, Perú, taxonomía Abstract The family Acanthaceae in Peru has just been reviewed by Brako and Zarruchi in 1993, since then, new information about this family has been generated. The present work is a synopsis of family Acanthaceae where four subfamilies (includying Avicennioideae) and 38 genera are recognized. -

11-FLOWER DIAGRAMES, FORMULAS and FLOWER SYMETRY FLOWER FORMULAS and DIAGRAMES
11-FLOWER DIAGRAMES, FORMULAS AND FLOWER SYMETRY FLOWER FORMULAS and DIAGRAMES 1. FLOWER FORMULAS Floral formula is a means to represent the structure of a flower using numbers, letters and various symbols, presenting substantial information about the flower in a compact form. It can represent particular species, or can be generalized to characterize higher taxa, usually giving ranges of organ numbers. Floral formulae are one of the two ways of describing flower structure developed during the 19th century, the other being floral diagrams. Apart from the graphical diagrams, the flower structure can be characterized by textual formulae. A floral formula consists of five symbols indicating from left to right: Floral Symmetry Number of Tepal Number of Sepals Number of Petals Number of Stamens Number of Carpels Tepals Sepals Patals Stamen Carpels P K C A G The parts of the flower are described according to their arrangement from the outside to the inside of the flower. If an organ type is arranged in more whorls, the outermost is denoted first, and the whorls are separated by “+”. If the organ number is large or fluctuating, is denoted as “∞”. 2. FLOWER DIAGRAMES Floral diagram is a graphic representation of flower structure. It shows the number of floral organs, their arrangement and fusion. Different parts of the flower are represented by their respective symbols. Rather like floral formulas, floral diagrams are used to show symmetry, numbers of parts, the relationships of the parts to one another, and degree of connation and/or adnation. Such diagrams cannot easily show ovary position. FLOWER SYMMETRY Floral symmetry describes whether, and how, a flower in particular its perianth, can be divided into two or more identical or mirror-image parts. -

Basic Botany
Basic Botany - Flower Structure The Birmingham Botanical Gardens & Glasshouses Brief Descriptions of Activities Flower Structure • a Study Centre-led activity • Using large-scale models and bee (glove puppet) to take pupils through the basic flower parts and their functions Investigating Floral Structure A wide range of flowers are always on display in the glasshouses. Their structure can be recorded in a variety of ways: • Directed observation through use of questionnaires • Drawing half a flower and labelling its structure • Creating a plan of the flower as if viewed from above • Creating a simple floral formula (this worksheet is using a simplified form of the recording system used by botanists) See worksheets 1-4 at back of booklet. Pollination Mechanisms • An extension of this work is to look at a variety of ways in which plants are designed in order to attract different pollinators See ‘A Guide To Pollinators’ at back of booklet. • Busy Bees. This is a game where pupils act out pollination See worksheet 5 at back of booklet Guide To Pollinators “Bee Flowers” Typically yellow, blue or purple. They produce pollen and lots of nectar, are often marked with lines and blotches and are sweetly scented at certain times of the day. “Butterfly Flowers” Vivid colours, often purple, red or white. Usually open during the day with a long thin corolla tube, lots of nectar and a strong scent. “Moth Flowers” Often white, p ink or pale yellow, open at night and have a heavy scent. “Wasp Flowers” Often pinkish or dirty red, with horizontal or drooping cups into which the short tongued wasp can push its head. -

Downloaded and Set As out Groups Genes
Alzahrani et al. BMC Genomics (2020) 21:393 https://doi.org/10.1186/s12864-020-06798-2 RESEARCH ARTICLE Open Access Complete chloroplast genome sequence of Barleria prionitis, comparative chloroplast genomics and phylogenetic relationships among Acanthoideae Dhafer A. Alzahrani1, Samaila S. Yaradua1,2*, Enas J. Albokhari1,3 and Abidina Abba1 Abstract Background: The plastome of medicinal and endangered species in Kingdom of Saudi Arabia, Barleria prionitis was sequenced. The plastome was compared with that of seven Acanthoideae species in order to describe the plastome, spot the microsatellite, assess the dissimilarities within the sampled plastomes and to infer their phylogenetic relationships. Results: The plastome of B. prionitis was 152,217 bp in length with Guanine-Cytosine and Adenine-Thymine content of 38.3 and 61.7% respectively. It is circular and quadripartite in structure and constitute of a large single copy (LSC, 83, 772 bp), small single copy (SSC, 17, 803 bp) and a pair of inverted repeat (IRa and IRb 25, 321 bp each). 131 genes were identified in the plastome out of which 113 are unique and 18 were repeated in IR region. The genome consists of 4 rRNA, 30 tRNA and 80 protein-coding genes. The analysis of long repeat showed all types of repeats were present in the plastome and palindromic has the highest frequency. A total number of 98 SSR were also identified of which mostly were mononucleotide Adenine-Thymine and are located at the non coding regions. Comparative genomic analysis among the plastomes revealed that the pair of the inverted repeat is more conserved than the single copy region. -

Sample Chapter
CHAPTER 2 Description of Plants PRINCIPLES OF PLANT DESCRIPTION HABIT: Natural locality of plants. • Ornamental plants: Plants cultivated for its beauty rather than its use. e.g. Marigold, Gladiolus etc. • Food crop: For economic use e.g. Maize, Rice, Apple etc. • Wild crop: Grow or produced without human care. e.g. Wild rice (Zizania aquatica), Wild rye (Elymus spp.). HABITAT: Place where a plant lives and grows. • Annual: Occurring every year. e.g. Rice, Brinjal etc. • Biennial: Occurring every two years. e.g. Raddish, Turnip etc. • Perennial: Present in all seasons of year i.e. continual. e.g. Mango, Rose etc. NATURE: Inherent or basic character. • Herb: Bushy, non-woody, erect, prostrate and decumbent. e.g. Mint, Hyacinth etc. • Shrub: Several stemmed, medium-sized woody plant. e.g. Jasmine, Rose etc. • Tree: Stout, woody trunk with few or no branches on its lower part, perennial. e.g. Mango, Pine, Banyan etc. • Clums: Nodes and internodes clearly visible. e.g. Bambusa These may be a) Deciduous—Falling off leaves annually. b) Evergreen—Having foliage leaves which remain green. c) Perennial—Persists for several years. Root Organ of a plant which grows downwards, away from light and towards water. It doesn’t bear leaves and buds but has protective apex called root cap. 10 Introduction to Pharmacognosy • Assimilatory root: Roots become green and serve for photosynthesis. e.g. Trapa • Tuberous root: Swollen, root without any definite shape. e.g. Sweet potato • Fasciculated root: Several tuberous roots occur in cluster at the base of stem. e.g. Dahlia • Nodulous root: Tuberous root becomes suddenly swollen at apex. -

Mimusops Elengi Linn:A Review
Review article Rapports De Pharmacie Vol.2 (3), 2016, 270-278 ISSN: 2455-0507 MIMUSOPS ELENGI LINN:A REVIEW Anuradha.S.N1, Arunkumar.S2 1Unit of Pharmaceutical Technology, Faculty of Pharmacy, Asian Institute of Medicine, Science and Technology (AIMST) University, Bedong 08100, Kedah, Malaysia 2Scientist, IPDO, Dr.reddys lab, Hyderabad, India. ABSTRACT Malaysia has diverse population combined by three races with their traditional system of medications. Large fraction of this population still depends mainly on traditional medicine. Use of herbal products is getting familiar in the growing generation as awareness on personnel healthcare has become a way of life. The naturally available plant products proved to be a no side effect medication foe a numerous diseases conditions. One of the traditional Indian system of medicine, Siddha uses the bark, fruit and seeds of Mimusops Elengi, which has several medicinal properties. Various plant parts(root, bark, leaves, flowers, fruit & seeds) have been found to be useful as cardio tonic, alexipharmic, stomachic, hypotensive, antibacterial, anthelmintic, antiulcer, teeth strengtheners and renewable source of energy. Keywords: Mimusops Elengi Linn, Phytoconstituents, Pharmacological Activites INTRODUCTION Traditional medicinesare being practiced from the researches on the medicinal plants about their early human civilizations. Based on hundreds of medicinal values have forced scientists to search for years of belief, observations, and analysis the plant-derived drugs for treatment of illness. pharmacological action of these medicines were Therefore, there has been growing interest among established. Different parts of the plant have scientists to isolate and study the pharmacological different pharmacological actions, which isevaluated properties of the phytochemicals. Herbal medicines and used in development of modern medicine. -

Agneta Julia Borg
Evolutionary relationships in Thunbergioideae and other early branching lineages of Acanthac e a e Agneta Julia Borg Evolutionary relationships in Thunbergioideae and other early branching lineages of Acanthaceae Agneta Julia Borg ©Agneta Julia Borg, Stockholm 2012 Cover illustration: From top left, Mendoncia retusa, Thunbergia convolvulifolia , Pseudocalyx saccatus, Crossandra strobilifera, Avicennia bicolor, Elytraria marginata. Photos: Agneta Julia Borg and Jürg Schönenberger. ISBN 978-91-7447-445-9 Printed in Sweden by Universitetsservice US-AB, Stockholm 2012 Distributor: Department of Botany, Stockholm University Academic dissertation for the degree of Doctor of Philosophy in Plant Sys- tematics presented at Stockholm University 2012 Abstract Borg, A.J. 2012. Evolutionary relationships in Thunbergioideae and other early branching lineages of Acanthaceae. Acanthaceae as circumscribed today consists of the three subfamilies Acan- thoideae (Acanthaceae sensu stricto), Thunbergioideae and Nelsonioideae, plus the genus Avicennia. Due to the morphological dissimilarities of Thun- bergioideae and Nelsonioideae, the delimitation of the family has been con- troversial. The mangrove genus Avicennia was only recently associated with Acanthaceae for the first time, based on molecular evidence, but without morphological support. In this thesis, phylogenetic analyses of nuclear and chloroplast DNA sequences were used to test the monophyly and exact posi- tions of Thunbergioideae and Nelsonioideae, and to infer detailed phyloge- netic relationships within these subfamilies and among major lineages of Acanthaceae. Floral structure and development were comparatively studied in Avicennia and other Acanthaceae using scanning electron microscopy and stereo microscopy. Phylogenetic analyses strongly support monophyly of Thunbergioideae and Nelsonioideae, and place the latter clade with strong support as sister to all other plants treated as Acanthaceae.