Araneae, Philodromidae)
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

SPIDERS of WASHINGTON COUNTY, MISSISSIPPI Orrey P. Young Southern Field Crop Insect Management Laboratory USDA-ARS, P.O. Box
Young, O . P., T. C . Lockley and G . B . Edwards . 1989 . Spiders of Washington County, Mississippi . J . Arachnol ., 17 :27-41 . SPIDERS OF WASHINGTON COUNTY, MISSISSIPPI Orrey P. Young Southern Field Crop Insect Management Laboratory USDA-ARS, P.O. Box 346 Stoneville, Mississippi 38776 USA Timothy C. Lockley Imported Fire Ant Station USDA-APHIS-PPQ 3505 25th Avenue Gulfport, Mississippi 39501 USA and G. B. Edwards Florida State Collection of Arthropods Division of Plant Industry Florida Dept. Agric. & Cons . Serv. P.O. Box 1269 Gainesville, Florida 32602 USA ABSTRACT Over a seven-year period, approximately 35,000 spiders representing 26 families, 133 genera, and 234 species were captured in Washington County, Mississippi, by pitfall, sweepnet, vacuum, bag, and hand. Specimens were collected in 10 different habitat types and in four vegetational strata . Old-field habitats yielded the most species (152) and residential lawns the fewest (14) . Considering all habitats sampled, the ground layer produced 111 species, the herbaceous strata 133, the shrub layer 49, and the tree strata 30 species . The sweepnet method of capture obtained 128 species, pitfall 95, hand 61, vacuum 53, and bagging 19 species. The largest number of species were obtained in spring and early summer (maximum of 125 in May), with the fewest in mid-winter (Jan . = 24) . Twenty-one species were considered abundant, 51 common, 67 uncommon, and 95 rare . Additions to the state list of Dorris (1972) number 102 species, for a new state total of 364 species . A comparison with the North American fauna and with other surveys indicates that Washington County is underrepresented both in cursorial forms active on the soil surface and web-spinning forms typical of undisturbed habitats . -
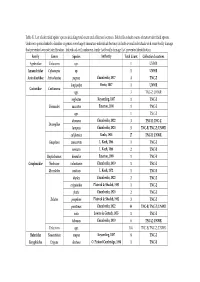
Table S1. List of Identified Spider Species Including Total Count and Collection Locations. Bold Cells Include Counts of Mature Identified Species
Table S1. List of identified spider species including total count and collection locations. Bold cells include counts of mature identified species. Unknown species linked to families or genera were largely immature individuals but may include several individuals with some bodily damage that prevented accurate identification. Individuals with unknown family had bodily damage that prevented identification. Family Genus Species Authority Total Count Collection Locations Agelenidae Unknown spp. 1 UNWR Amaurobiidae Cybaeopsis sp. 1 UNWR Antrodiaetidae Antrodiaetus pugnax Chamberlin, 1917 4 TNC-Z longipalpa Hentz, 1847 1 UNWR Corinnidae Castianeira spp. 3 TNC-Z; UNWR neglectus Keyserling, 1887 1 TNC-Z Drassodes saccatus Emerton, 1890 1 TNC-Z spp. 1 TNC-Z dromeus Chamberlin, 1922 3 TNC-B; TNC-Z Drassyllus lamprus Chamberlin, 1920 3 TNC-B; TNC-Z; UNWR californica Banks, 1904 17 TNC-B; UNWR Gnaphosa muscorum L. Koch, 1866 1 TNC-Z sericata L. Koch, 1866 2 TNC-B Haplodrassus hiemalis Emerton, 1909 1 TNC-B Gnaphosidae Nodocion voluntaries Chamberlin, 1919 1 TNC-Z Urozelotes rusticus L. Koch, 1872 1 TNC-B duplex Chamberlin, 1922 2 TNC-Z exiguioides Platnick & Shadab, 1983 1 TNC-Z fratis Chamberlin, 1920 2 TNC-Z Zelotes josephine Platnick & Shadab, 1983 3 TNC-Z puritanus Chamberlin, 1922 44 TNC-B; TNC-Z; UNWR sula Lowrie & Gertsch, 1955 1 TNC-Z tubuous Chamberlin, 1919 6 TNC-Z; UNWR Unknown spp. 184 TNC-B; TNC-Z; UNWR Hahniidae Neoantistea magna Keyserling, 1887 8 TNC-Z Linyphiidae Erigone dentosa O. Pickard-Cambridge, 1894 1 TNC-B spp. 1 TNC-Z Unknown spp. 13 TNC-Z; UNWR mccooki Montgomery, 1904 95 TNC-B; TNC-Z; UNWR Schizocosa minnesotensis Gertsch, 1934 25 TNC-B Lycosidae spp. -

SA Spider Checklist
REVIEW ZOOS' PRINT JOURNAL 22(2): 2551-2597 CHECKLIST OF SPIDERS (ARACHNIDA: ARANEAE) OF SOUTH ASIA INCLUDING THE 2006 UPDATE OF INDIAN SPIDER CHECKLIST Manju Siliwal 1 and Sanjay Molur 2,3 1,2 Wildlife Information & Liaison Development (WILD) Society, 3 Zoo Outreach Organisation (ZOO) 29-1, Bharathi Colony, Peelamedu, Coimbatore, Tamil Nadu 641004, India Email: 1 [email protected]; 3 [email protected] ABSTRACT Thesaurus, (Vol. 1) in 1734 (Smith, 2001). Most of the spiders After one year since publication of the Indian Checklist, this is described during the British period from South Asia were by an attempt to provide a comprehensive checklist of spiders of foreigners based on the specimens deposited in different South Asia with eight countries - Afghanistan, Bangladesh, Bhutan, India, Maldives, Nepal, Pakistan and Sri Lanka. The European Museums. Indian checklist is also updated for 2006. The South Asian While the Indian checklist (Siliwal et al., 2005) is more spider list is also compiled following The World Spider Catalog accurate, the South Asian spider checklist is not critically by Platnick and other peer-reviewed publications since the last scrutinized due to lack of complete literature, but it gives an update. In total, 2299 species of spiders in 67 families have overview of species found in various South Asian countries, been reported from South Asia. There are 39 species included in this regions checklist that are not listed in the World Catalog gives the endemism of species and forms a basis for careful of Spiders. Taxonomic verification is recommended for 51 species. and participatory work by arachnologists in the region. -

Araneae : Philodromidae and Thomisidae, by Charles D
The Journal of Arachnology 8 : 95 Muchmore, W . B. 1969a. The pseudoscorpion genus Neochthonius Chamberlin (Arachnida , Chelonethida, Chthoniidae) with description of a cavernicolous species, Amer. Midl . Nat . 81 :387-394 . Muchmore, W. B . 1969b . New species and records of cavernicolous pseudoscorpions of the genu s Microcreagris (Arachnida, Chelonethida, Neobisiidae, Ideobisiinae) . Amer . Mus. Novitate s 2392:1-21 . Muchmore, W. B . 1976 . New species of Apochthonius, mainly from caves in central and eastern United States (Pseudoscirpionida, Chthoniidae) . Proc. Biol . Soc . Washington 89 :67-80 . Muchmore, W . B. and E. M . Benedict . 1976 . Redescription of Apochthonius moestus (Banks), type of the genus Apochthonius Chamberlin (Pseudoscirpionida, Chthoniidae) . J. New York Entomol. Soc. 84 :67-74 . Schuster, R . O . 1966 . New species of Apochthonius from western North America . Pan-Pacifi c Entomol . 42 :178-183 . Manuscript received March 1979, revised May 1979. BOOK REVIEW The Crab Spiders of Canada and Alaska : Araneae : Philodromidae and Thomisidae, by Charles D . Dondale and James H. Redner, 1978 . Part 5 of The Insects and Arachnida o f Canada, Publication 1663, pp . 1-255, 725 figs., 66 maps . Available from Printing and Publishing Supply and Services Canada, Hull, Quebec, Canada K1A OS9, Canada . Canada : $7 .50 ; other countries : $9.00. This handsome, soft-bound volume, dealing with the spider families Philodromidae an d Thomisidae from north of the United States, is a striking contribution to Canadia n arachnology . About half of the taxa of all temperate North America (110 of some 22 3 species) occur in Canada . The authors have drawn much of the data from their many papers on these spiders, but this has been materially supplemented by new appraisals , illustrations, and distribution information . -

Genetic Diversity Analysis of Crab Spider (Araneae: Thomisidae) Based on RAPD-PCR
RESEARCH PAPER Zoology Volume : 4 | Issue : 8 | August 2014 | ISSN - 2249-555X Genetic Diversity Analysis of Crab Spider (Araneae: Thomisidae) based on RAPD-PCR KEYWORDS Crab spiders, RAPD, diversity, Amravati Nandkishor Warghat Navin Sharma Assistant professor, Department of Zoology, Arts, Assistant professor, Department of Zoology, Arts and commerce and Science College , Maregaon, Dist- Science College, Pulgaon, Dist- Wardha, Maharashtra. Yavatmal, Maharashtra. Punam Thakur *Mumtaz Baig Associate professor, Department of Zoology, Govt. Research Scholar, Department of Zoology, Govt. Vidarbha Institute of Science and Humanities, Vidarbha Institute of Science and Humanities, Amravati Amravati-444604. MH *Corresponding Author ABSTRACT The current study deals with the genetic diversity of crab spiders using molecular markers. Total scorable bands were produced using six random primers for the 16 species of crab spiders belonging to family thomisidae. Out of all screened primers, OPP 9 produced highest 113 scorable bands, out of which 111 bands were found with 98.28 percent polymorphism. Primer OPA 3 was produced lowest 43 bands with 100 percent polymor- phism. Remaining primers (OPA 2, OPA 4, OPN 16 and OPN 18) showed 100 per cent polymorphism with 60, 60, 81 and 51 bands respectively. Along with the molecular markers, a morphometric analysis was also focus on the phyloge- netic relationship of crab spiders using UPGMA and NJ approach. The present study is the first report from India to describe the genetic relatedness amongst spider using RAPD-PCR. Introduction at 20055’ and 20.93 North latitude 77045’ and 77.75 East Crab spiders are small to large size, with two claws and longitudes at an elevation of 1125 feet. -

Special Publications Special
ARACHNIDS ASSOCIATED WITH WET PLAYAS IN THE SOUTHERN HIGH PLAINS WITH WET PLAYAS ARACHNIDS ASSOCIATED SPECIAL PUBLICATIONS Museum of Texas Tech University Number 54 2008 ARACHNIDS ASSOCIATED WITH WET PLAYAS IN THE SOUTHERN HIGH PLAINS (LLANO ESTACADO), C okendolpher et al. U.S.A. JAMES C. COKENDOLPHER, SHANNON M. TORRENCE, JAMES T. ANDERSON, W. DAVID SISSOM, NADINE DUPÉRRÉ, JAMES D. RAY & LOREN M. SMITH SPECIAL PUBLICATIONS Museum of Texas Tech University Number 54 Arachnids Associated with Wet Playas in the Southern High Plains (Llano Estacado), U.S.A. JAMES C. COKENDOLPHER , SHANNON M. TORREN C E , JAMES T. ANDERSON , W. DAVID SISSOM , NADINE DUPÉRRÉ , JAMES D. RAY , AND LOREN M. SMI T H Texas Tech University, Oklahoma State University, B&W Pantex, Texas Parks and Wildlife Department, West Texas A&M University, West Virginia University Layout and Design: Lisa Bradley Cover Design: James C. Cokendolpher et al. Copyright 2008, Museum of Texas Tech University All rights reserved. No portion of this book may be reproduced in any form or by any means, including electronic storage and retrieval systems, except by explicit, prior written permission of the publisher. This book was set in Times New Roman and printed on acid-free paper that meets the guidelines for permanence and durability of the Committee on Production Guidelines for Book Longevity of the Council on Library Resources. Printed: 10 April 2008 Library of Congress Cataloging-in-Publication Data Special Publications of the Museum of Texas Tech University, Number 54 Series Editor: Robert J. Baker Arachnids Associated with Wet Playas in the Southern High Plains (Llano Estacado), U.S.A. -

University Morifilms International 300N.Zeeb Road Ann Arbor, Ml 48106 8305402
INFORMATION TO USERS This reproduction was made from a copy of a document sent to us for microfilming. While the most advanced technology has been used to photograph and reproduce this document, the quality of the reproduction is heavily dependent upon the quality of the material submitted. The following explanation of techniques is provided to help clarify markings or notations which may appear on this reproduction. 1.The sign or “target” for pages apparently lacking from the document photographed is “Missing Page(s)”. If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting through an image and duplicating adjacent pages to assure complete continuity. 2. When an image on the film is obliterated with a round black mark, it is an indication of either blurred copy because of movement during exposure, duplicate copy, or copyrighted materials that should not have been filmed. For blurred pages, a good image of the page can be found in the adjacent frame. If copyrighted materials were deleted, a target note will appear listing the pages in the adjacent frame. 3. When a map, drawing or chart, etc., is part of the material being photographed, a definite method of “sectioning” the material has been followed. It is customary to begin filming at the upper left hand comer of a large sheet and to continue from left to right in equal sections with small overlaps. If necessary, sectioning is continued again-beginning below the first row and continuing on until complete. -

1 CHECKLIST of ILLINOIS SPIDERS Over 500 Spider Species Have Been
1 CHECKLIST OF ILLINOIS SPIDERS Over 500 spider species have been reported to occur in Illinois. This checklist includes 558 species, and there may be records in the literature that have eluded the author’s attention. This checklist of Illinois species has been compiled from sources cited below. The initials in parentheses that follow each species name and authorship in the list denote the paper or other source in which the species was reported. Locality data, dates of collection, and other information about each species can be obtained by referring to the indicated sources. (AAS) American Arachnological Society Spider Species List for North America, published on the web site of the American Arachnological Society: http://americanarachnology.org/AAS_information.html (B&N) Beatty, J. A. and J. M. Nelson. 1979. Additions to the Checklist of Illinois Spiders. The Great Lakes Entomologist 12:49-56. (JB) Beatty, J. A. 2002. The Spiders of Illinois and Indiana, their Geolographical Affinities, and an Annotated Checklist. Proc. Ind. Acad. Sci. 1:77-94. (BC) Cutler, B. 1987. A Revision of the American Species of the Antlike Jumping Spider Genus Synageles (Araneae: Salticidae). J. Arachnol.15:321-348. (G&P) Gertsch, W. J. And N. I. Platnick. 1980. A Revision of the American Spiders of the Family Atypidae (Araneae, Mygalomorphae). Amer. Mus. Novitates 2704:1-39. (BK) Kaston, B. J. 1955. Check List of Illinois Spiders. Trans. Ill. State Acad. Sci. 47: 165- 172. (SK) Kendeigh, S. C. 1979. Invertebrate Populations of the Deciduous Forest Fluctuations and Relations to Weather. Illinois Biol. Monog. 50:1-107. -

Spiders of Alberta: from Agelenidae to Uloboridae
Spiders of Alberta: from Agelenidae to Uloboridae Dr. Heather Proctor University of Alberta for the Edmonton Nature Club, 7 Feb 2019 (selected slides for posting; photos (c) H. Proctor unless otherwise noted) Canadian and Albertan diversity • 1477 species of spiders in 45 families known from Canada – may be up to 1800 spp. • 657 species in 28 families known from Alberta 631 of the 657 species are included here from https://www.albertaparks.ca/media/6255191/list-of-elements-ab-invertebrates-spiders.xlsx The 28 families of spiders known from Alberta • no mygalomorph spiders in AB, only araneomorph • Division Synspermiata – Pholcioidea: Pholcidae, Telemidae • Division Entelegynae – Araneoidea: Theridiidae, Araneidae, Linyphiidae, Mysmenidae, Mimetidae, Tetragnathidae – Uloboroidea: Uloboridae – Titanoecoidea: Titanoecidae – Amaurobioidea: Amaurobiidae – Desoidea: Desidae – Agelenoidea: Dictynidae, Cybaeidae, Hahniidae, Agelenidae – Lycosoidea: Oxyopidae, Thomisidae, Pisauridae, Lycosidae – Salticoidea: Salticidae, Philodromidae, Corinnidae, Eutichuridae – Anyphaenoidea: Anyphaenidae, Clubionidae – Liocranoidea: Liocranidae – Trochanteroidea: Phrurolithidae, Gnaphosidae mygalomorphs from BC, Antrodiaetus sp. Linyphiidae 261 Gnaphosidae 51 Lycosidae 50 Salticidae 45 Number of species known Dictynidae 36 from each family in Alberta Thomisidae 37 Theridiidae 36 (based on Robb Bennett’s Araneidae 32 personal list, 7 Feb 2019) Philodromidae 29 Clubionidae 17 Tetragnathidae 14 Hahniidae 10 Amaurobiidae 7 Agelenidae 6 Corinnidae 3 Phrurolithidae -

Research Notes
Arthropod Community Dynamics in New Varietal Vineyards CATI Publication No. 9810G3 Viticulture and Enology Research Center October 1998 CONCLUSIONS spider Ebo pepinensis,as well as various species of General summary observations from the 1997 wingless scelionid and ichneumonid wasps. Ripperdan Project include the following: 3) Densities of important insect pests, such as 1) Spiders and beneficialinsects can be quite abundant Erythroneura leafhoppers and omnivorous leaf-roller larvae, may remain very low throughout an entire Research Notes in established, well-balanced vineyard ecosystems, growing season coincident with natural enemy but they may also reach relatively high densities in numbers at relatively high levels. For example, field non-insecticide treated vineyards during the first- parasitization rates for OLR may at times reach 50% California State University, Fresno leaf growth season. (far above the more commonly reported 10% level). 2) High numbers of relatively unknown natural 4) From the relatively limited results of the first season enemies may be found in typical San Joaquin Valley Arthropod Community Dynamics In New of this ongoing research project, no evidence was vineyards when appropriate care is given to sam Varietal Vineyards pling techniques and procedures. Examples of these collected which suggests any particular arthropod obscure predators and parasitoids include the pest problems in the less traditional grape varietals being planted in the San Joaquin Valley. by Mark A. Mayse, Jo Harper and Jason Stock previously umeported vineyard philodromid INTRODUCTION fumigated with methyl bromide, and the remaining During recent years, a number of San Joaquin Valley 6-acre portion left non-fumigated; and 2) a lOO-acre winegrape growers have decided to investigate the Chardonnay block (791CH, comprised of 64 acres About CATI.. -

Diversity and Phenology of Predatory Arthropods Overwintering in Cardboard Bands Placed in Pear and Apple Orchards of Central Washington State
ECOLOGY AND POPULATION BIOLOGY Diversity and Phenology of Predatory Arthropods Overwintering in Cardboard Bands Placed in Pear and Apple Orchards of Central Washington State DAVID R. HORTON,1 DEBRA A. BROERS, TONYA HINOJOSA, TAMERA M. LEWIS, EUGENE R. MILICZKY, AND RICHARD R. LEWIS USDAÐARS, 5230 Konnowac Pass Road, Wapato, WA 98951 Ann. Entomol. Soc. Am. 95(4): 469Ð480 (2002) ABSTRACT Overwintering shelters composed of cardboard bands were placed on pear and apple trees located in central Washington state to monitor overwintering by predatory arthropods and by two pest taxa. A subset of bands was sampled at regular intervals between late summer and mid- December to determine when taxa began to enter bands. The remaining bands were left undisturbed until collection in mid-December to determine the numbers and types of arthropods overwintering on tree trunks in these orchards. More than 8,000 predatory arthropods were collected from bands left undisturbed until mid-December, dominated numerically by Acari (Phytoseiidae) [Galendromus occidentalis (Nesbitt), Typhlodromus spp.], Araneae, and Neuroptera (Hemerobiidae, Chrysopidae). Predatory mite numbers were higher in bands placed in apple orchards than bands placed in pear orchards. The Araneae were particularly diverse, including Ͼ3,000 spiders representing nine families. Less abundant were Heteroptera, including a mirid [Deraeocoris brevis (Uhler)] and three species of Anthocoridae [Anthocoris spp., Orius tristicolor (White)]. Coleoptera included Coccinellidae, dom- inated by Stethorus picipes Casey, and unidentiÞed Staphylinidae and Carabidae. The bands that were collected at regular intervals to monitor phenology provided Ͼ15,000 predatory arthropods, domi- nated numerically by spiders, Dermaptera [Forficula auricularia (F.)], lacewings, and predatory mites. -

Arachnida: Araneae) of the Canadian Prairies
75 Chapter 4 Spiders (Arachnida: Araneae) of the Canadian Prairies Héctor Cárcamo Lethbridge Research Centre, Agriculture and Agri-Food Canada, Lethbridge, AB Jaime Pinzón Department of Renewable Resources, Faculty of Agricultural, Life and Environmental Sciences, University of Alberta, Edmonton Robin Leech 10534, 139 St NW, Edmonton AB John Spence Department of Renewable Resources, Faculty of Agricultural, Life and Environmental Sciences, University of Alberta, Edmonton Abstract. Spiders are the seventh most diverse order of arthropods globally and are prominent predators in all prairie habitats. In this chapter, a checklist for the spiders of the Prairie Provinces (767 recorded species and 44 possible species) is presented along with an overview of all 26 families that occur in the region. Eighteen of the species from the region are adventive. Linyphiidae is by far the dominant family, representing 39% of all species in the three provinces. Gnaphosidae and Lycosidae each represent 8% and three other families (Salticidae, Dictynidae, and Theridiidae) each account for 7%. A summary of biodiversity studies conducted in the Prairies Ecozone and from transition ecoregions is also provided. The Mixed Grassland Ecoregion has the most distinctive assemblage; Schizocosa mccooki and Zelotes lasalanus are common only in this ecoregion. Other ecoregions appear to harbour less distinctive assemblages, but most have been poorly studied. Lack of professional opportunities for spider systematists in Canada remains a major barrier to the advancement of the taxonomy and ecology of spiders. Résumé. Les aranéides forment le septième ordre le plus diversifi é d’arthropodes dans le monde; ce sont des prédateurs très présents dans tous les habitats des Prairies.