Diversity and Phenology of Predatory Arthropods Overwintering in Cardboard Bands Placed in Pear and Apple Orchards of Central Washington State
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Green Lacewings of the Genus Chrysopa in Maryland ( Neuroptera: Chrysopidae)
The Green Lacewings of the Genus Chrysopa in Maryland ( Neuroptera: Chrysopidae) Ralph A. Bram and William E. Bickley Department of Entomology INTRODUCTION Tlw green lacewings which are members of the genus Chrysopa are extreme- ly lwndicia1 insects. The larvae are commonly called aphislions and are well known as predators of aphids and other injurious insects. They play an important part in the regulation of populations of pests under natural conditions, and in California they have been cultured in mass and released for the control of mealy- bugs ( Finney, 1948 and 1950) . The positive identification of members of the genus is desirable for the use of biological-control workers and entomologists in general. Descriptions of most of the Nearctic species of Chrysopidae have relied heavily on body pigmentation and to a lesser extent on wing shape, venational patterns and coloration. Specimens fade when preserved in alcohol or on pins, and natural variation in color patterns occurs in many species ( Smith 1922, Bickley 1952). It is partly for these reasons that some of the most common and relatively abundant representatives of the family are not easily recognized. The chrysopid fauna of North America was treated comprehensively by Banks ( 1903). Smith ( 1922) contributed valuable information about the biology of the green lacewings and about the morphology and taxonomy of the larvae. He also pro- vided k<'ys and other help for the identification of species from Kansas ( 1925, 1934) and Canada ( 1932). Froeschner ( 194 7) similarly dealt with Missouri species. Bickley and MacLeod ( 1956) presented a review of the family as known to occur in the N earctic region north of Mexico. -

Monitoring of Pear Psylla for Pest Management Decisions and Research
Integrated Pest Management Reviews 4: 1–20, 1999. © 1999 Kluwer Academic Publishers. Printed in the Netherlands. Monitoring of pear psylla for pest management decisions and research David R. Horton USDA-ARS, 5230 Konnowac Pass Rd., Wapato, WA 98951 U.S.A. (Tel.: (509) 454-5639; Fax: 509-454-5646) Received 6 May 1998; accepted 3 November 1998 Abstract Pear psylla, Cacopsylla pyricola (Foerster) (Homoptera: Psyllidae), is one of the key insect pests in North American pear production. In some growing areas, more than 50% of dollars spent to control arthropod pests in commercial pear are directed specifically at controlling this species. Control measures require accurate and timely information about dispersal, onset of egg-laying in spring, densities in the orchard, and age composition of the population. To meet these ends, a number of sampling methods have been developed to monitor pear psylla, the most common being (for pest management purposes) visual inspection of spurs and foliage for nymphs and eggs, and use of beat trays to monitor adults. Action thresholds have been developed for counts obtained with either method. However, threshold estimates are fairly narrowly defined, referring to a somewhat limited group of pear cultivars, type of injury to be prevented, and pest management program being used. Further refinement has been difficult because of an incomplete understanding of psylla’s spatial distribution, seasonal changes in spatial distribution, and unknown or seasonally changing action thresholds. Beat trays and visual inspection of foliage have also been used to monitor pear psylla in various types of research projects, including studies of dispersal and biological control. -

Distribution of Spiders in Coastal Grey Dunes
kaft_def 7/8/04 11:22 AM Pagina 1 SPATIAL PATTERNS AND EVOLUTIONARY D ISTRIBUTION OF SPIDERS IN COASTAL GREY DUNES Distribution of spiders in coastal grey dunes SPATIAL PATTERNS AND EVOLUTIONARY- ECOLOGICAL IMPORTANCE OF DISPERSAL - ECOLOGICAL IMPORTANCE OF DISPERSAL Dries Bonte Dispersal is crucial in structuring species distribution, population structure and species ranges at large geographical scales or within local patchily distributed populations. The knowledge of dispersal evolution, motivation, its effect on metapopulation dynamics and species distribution at multiple scales is poorly understood and many questions remain unsolved or require empirical verification. In this thesis we contribute to the knowledge of dispersal, by studying both ecological and evolutionary aspects of spider dispersal in fragmented grey dunes. Studies were performed at the individual, population and assemblage level and indicate that behavioural traits narrowly linked to dispersal, con- siderably show [adaptive] variation in function of habitat quality and geometry. Dispersal also determines spider distribution patterns and metapopulation dynamics. Consequently, our results stress the need to integrate knowledge on behavioural ecology within the study of ecological landscapes. / Promotor: Prof. Dr. Eckhart Kuijken [Ghent University & Institute of Nature Dries Bonte Conservation] Co-promotor: Prf. Dr. Jean-Pierre Maelfait [Ghent University & Institute of Nature Conservation] and Prof. Dr. Luc lens [Ghent University] Date of public defence: 6 February 2004 [Ghent University] Universiteit Gent Faculteit Wetenschappen Academiejaar 2003-2004 Distribution of spiders in coastal grey dunes: spatial patterns and evolutionary-ecological importance of dispersal Verspreiding van spinnen in grijze kustduinen: ruimtelijke patronen en evolutionair-ecologisch belang van dispersie door Dries Bonte Thesis submitted in fulfilment of the requirements for the degree of Doctor [Ph.D.] in Sciences Proefschrift voorgedragen tot het bekomen van de graad van Doctor in de Wetenschappen Promotor: Prof. -

A Summary List of Fossil Spiders
A summary list of fossil spiders compiled by Jason A. Dunlop (Berlin), David Penney (Manchester) & Denise Jekel (Berlin) Suggested citation: Dunlop, J. A., Penney, D. & Jekel, D. 2010. A summary list of fossil spiders. In Platnick, N. I. (ed.) The world spider catalog, version 10.5. American Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/index.html Last udated: 10.12.2009 INTRODUCTION Fossil spiders have not been fully cataloged since Bonnet’s Bibliographia Araneorum and are not included in the current Catalog. Since Bonnet’s time there has been considerable progress in our understanding of the spider fossil record and numerous new taxa have been described. As part of a larger project to catalog the diversity of fossil arachnids and their relatives, our aim here is to offer a summary list of the known fossil spiders in their current systematic position; as a first step towards the eventual goal of combining fossil and Recent data within a single arachnological resource. To integrate our data as smoothly as possible with standards used for living spiders, our list follows the names and sequence of families adopted in the Catalog. For this reason some of the family groupings proposed in Wunderlich’s (2004, 2008) monographs of amber and copal spiders are not reflected here, and we encourage the reader to consult these studies for details and alternative opinions. Extinct families have been inserted in the position which we hope best reflects their probable affinities. Genus and species names were compiled from established lists and cross-referenced against the primary literature. -

The Biology of the Predator Complex of the Filbert Aphid, Myzocallis Coryli
AN ABSTRACT OF THE THESIS OF Russell H. Messing for the degree of Master of Science in Entomology presented in July 1982 Title: The Biology of the Predator Complex of the Filbert Aphid, Myzocallis coryli (Goetze) in Western Oregon. Abstract approved: Redacted for Privacy M. T. AliNiiee Commercial filbert orchards throughout the Willamette Valley were surveyed for natural enemies of the filbert aphid, Myzocallis coryli (Goetze). A large number of predaceous insects were found to prey upon M. coryli, particularly members of the families Coccinellidae, Miridae, Chrysopidae, Hemerobiidae, and Syrphidae. Also, a parasitic Hymenopteran (Mesidiopsis sp.) and a fungal pathogen (Triplosporium fresenii) were found to attack this aphid species. Populations of major predators were monitored closely during 1981 to determine phenology and synchrony with aphid populations and to determine their relative importance. Adalia bipunctata, Deraeocoris brevis, Chrysopa sp. and Hemerobius sp. were found to be extremely well synchronized with aphid population development cycles. Laboratory feeding trials demonstrated that all 4 predaceous insects tested (Deraeocoris brevis, Heterotoma meriopterum, Compsidolon salicellum and Adalia bipunctata) had a severe impact upon filbert aphid population growth. A. bipunctata was more voracious than the other 3 species, but could not live as long in the absence of aphid prey. Several insecticides were tested both in the laboratory and field to determine their relative toxicity to filbert aphids and the major natural enemies. Field tests showed Metasystox-R to be the most effective against filbert aphids, while Diazinon, Systox, Zolone, and Thiodan were moderately effective. Sevin was relatively ineffective. All insecticides tested in the field severely disrupted the predator complex. -

Oregon Invasive Species Action Plan
Oregon Invasive Species Action Plan June 2005 Martin Nugent, Chair Wildlife Diversity Coordinator Oregon Department of Fish & Wildlife PO Box 59 Portland, OR 97207 (503) 872-5260 x5346 FAX: (503) 872-5269 [email protected] Kev Alexanian Dan Hilburn Sam Chan Bill Reynolds Suzanne Cudd Eric Schwamberger Risa Demasi Mark Systma Chris Guntermann Mandy Tu Randy Henry 7/15/05 Table of Contents Chapter 1........................................................................................................................3 Introduction ..................................................................................................................................... 3 What’s Going On?........................................................................................................................................ 3 Oregon Examples......................................................................................................................................... 5 Goal............................................................................................................................................................... 6 Invasive Species Council................................................................................................................. 6 Statute ........................................................................................................................................................... 6 Functions ..................................................................................................................................................... -
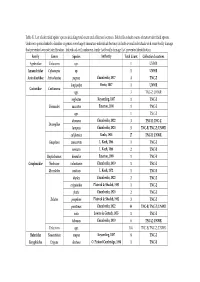
Table S1. List of Identified Spider Species Including Total Count and Collection Locations. Bold Cells Include Counts of Mature Identified Species
Table S1. List of identified spider species including total count and collection locations. Bold cells include counts of mature identified species. Unknown species linked to families or genera were largely immature individuals but may include several individuals with some bodily damage that prevented accurate identification. Individuals with unknown family had bodily damage that prevented identification. Family Genus Species Authority Total Count Collection Locations Agelenidae Unknown spp. 1 UNWR Amaurobiidae Cybaeopsis sp. 1 UNWR Antrodiaetidae Antrodiaetus pugnax Chamberlin, 1917 4 TNC-Z longipalpa Hentz, 1847 1 UNWR Corinnidae Castianeira spp. 3 TNC-Z; UNWR neglectus Keyserling, 1887 1 TNC-Z Drassodes saccatus Emerton, 1890 1 TNC-Z spp. 1 TNC-Z dromeus Chamberlin, 1922 3 TNC-B; TNC-Z Drassyllus lamprus Chamberlin, 1920 3 TNC-B; TNC-Z; UNWR californica Banks, 1904 17 TNC-B; UNWR Gnaphosa muscorum L. Koch, 1866 1 TNC-Z sericata L. Koch, 1866 2 TNC-B Haplodrassus hiemalis Emerton, 1909 1 TNC-B Gnaphosidae Nodocion voluntaries Chamberlin, 1919 1 TNC-Z Urozelotes rusticus L. Koch, 1872 1 TNC-B duplex Chamberlin, 1922 2 TNC-Z exiguioides Platnick & Shadab, 1983 1 TNC-Z fratis Chamberlin, 1920 2 TNC-Z Zelotes josephine Platnick & Shadab, 1983 3 TNC-Z puritanus Chamberlin, 1922 44 TNC-B; TNC-Z; UNWR sula Lowrie & Gertsch, 1955 1 TNC-Z tubuous Chamberlin, 1919 6 TNC-Z; UNWR Unknown spp. 184 TNC-B; TNC-Z; UNWR Hahniidae Neoantistea magna Keyserling, 1887 8 TNC-Z Linyphiidae Erigone dentosa O. Pickard-Cambridge, 1894 1 TNC-B spp. 1 TNC-Z Unknown spp. 13 TNC-Z; UNWR mccooki Montgomery, 1904 95 TNC-B; TNC-Z; UNWR Schizocosa minnesotensis Gertsch, 1934 25 TNC-B Lycosidae spp. -

Psyllid Host-Plants (Hemiptera: Psylloidea): Resolving a Semantic Problem
242 Florida Entomologist 97(1) March 2014 PSYLLID HOST-PLANTS (HEMIPTERA: PSYLLOIDEA): RESOLVING A SEMANTIC PROBLEM 1,* 2 3 2 DANIEL BURCKHARDT , DAVID OUVRARD , DALVA QUEIROZ AND DIANA PERCY 1Naturhistorisches Museum, Augustinergasse 2, CH-4001 Basel, Switzerland 2Department of Life Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, UK 3Embrapa Florestas, Colombo/PR, Brazil *Corresponding author; E-mail: [email protected] ABSTRACT Evolutionary and biological patterns can be obscured by inadequate or ill-defined terminol- ogy. An example is the generally very specific relationship between the sap-feeding hemip- teran group, psyllids, and their breeding plants, commonly called host-plants. The literature is clogged with references to so called ‘hosts’, which are often merely plants on which psyllids were found accidentally, and no immature development was detected. Recently the term host has also been applied by some authors to any plant on which immature or adults feed. Here we propose a terminology to clarify associated plant definitions, and we suggest restricting the use of the term host-plant to plants on which a psyllid species completes its immature to adult life cycle. For the other plant associations we suggest the terms overwintering or shel- ter plant (plants on which adult psyllids overwinter and on which they may feed), food plant (plants on which adult psyllids feed, but do not breed and do not spend an extended period of time) and casual plant (plants on which adult psyllids land but do not feed). Key Words: jumping plant-lice, psyllids, host-plant, terminology RESUMEN Patrones evolutivos y biológicos pueden ser oscurecidas por la terminología inadecuada o mal definida. -

Codling Moth Areawide Integrated Pest Management
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln U.S. Department of Agriculture: Agricultural Publications from USDA-ARS / UNL Faculty Research Service, Lincoln, Nebraska 2008 Codling Moth Areawide Integrated Pest Management Alan L. Knight USDA-ARS Follow this and additional works at: https://digitalcommons.unl.edu/usdaarsfacpub Part of the Agricultural Science Commons Knight, Alan L., "Codling Moth Areawide Integrated Pest Management" (2008). Publications from USDA- ARS / UNL Faculty. 651. https://digitalcommons.unl.edu/usdaarsfacpub/651 This Article is brought to you for free and open access by the U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Publications from USDA-ARS / UNL Faculty by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. - 9 Codling Moth Areawide Integrated Pest Management ALAN L. KNIGHT Yakima Agricultural Research Laboratory, Agricultural Research Service, Wapato, Washington, USA Introduction Codling moth (CM) is an insidious pest, tunnelling to the core of valuable commodi ties that are typically marketed with exceptional quality standards for appearance, firmness and sweetness. While there is no mention in the Bible of whether the apple that Eve gave to Adam graded 'Extra fancy', it is likely that if this fruit had been infested with CM, the human race would not be as anxious about returning to a pest-ridden garden of paradise. Nevertheless, since Noah allowed two adult CM to disembark from his boat, the distribution of this pest has closely followed man's culti vation of its hosts around the world (Shel'deshova, 1967). -

Impacts of Insecticides on Predatory Mite, Neoseiulus Fallacis (Acari: Phytoseidae) and Mite Flaring of European Red Mites, Panonychus Ulmi (Acari: Tetranychidae)
IMPACTS OF INSECTICIDES ON PREDATORY MITE, NEOSEIULUS FALLACIS (ACARI: PHYTOSEIDAE) AND MITE FLARING OF EUROPEAN RED MITES, PANONYCHUS ULMI (ACARI: TETRANYCHIDAE) By Raja Zalinda Raja Jamil A DISSERTATION Submitted to Michigan State University in partial fulfillment of the requirements for the degree of Entomology–Doctor of Philosophy 2014 ABSTRACT IMPACTS OF INSECTICIDES ON PREDATORY MITE, NEOSEIULUS FALLACIS (ACARI: PHYTOSEIDAE) AND MITE FLARING OF EUROPEAN RED MITES, PANONYCHUS ULMI (ACARI: TETRANYCHIDAE) By Raja Zalinda Raja Jamil Panonychus ulmi, the European red mite, is a major agricultural pest found in most deciduous fruit growing areas. It is the most important mite species attacking tree fruits in humid regions of North America. Bristle-like mouthparts of this mite species pierce the leaf cell wall and ingestion of their contents including chlorophyll causes bronzing injury to leaves. Heavy mite feeding early in the season (late Jun and July) reduce tree growth and yield as well as the fruit bud formation, thereby reduce yields the following year. Biological control of this pest species by predators has been a cornerstone of IPM. Phytoseiid mite, Neoseiulus fallacis (Garman) is the most effective predator mite in Michigan apple orchards and provides mid- and late-season biological control of European red mites. Achieving full potential of biological control in tree fruit has been challenging due to the periodic sprays of broad-spectrum insecticides. There have been cases of mite flaring reported by farmers in relation to the reduced-risk (RR) insecticides that were registered in commercial apple production in the past ten years. These insecticides are often used in fruit trees to control key direct pests such as the codling moth. -

Managing Floral Resources in Apple Orchards for Pest Control: Ideas, Experiences and Future Directions
insects Review Managing Floral Resources in Apple Orchards for Pest Control: Ideas, Experiences and Future Directions Annette Herz 1,*, Fabian Cahenzli 2, Servane Penvern 3, Lukas Pfiffner 2, Marco Tasin 4 and Lene Sigsgaard 5 1 Julius Kühn-Institut, Institute for Biological Control, Heinrichstr. 243, 64287 Darmstadt, Germany 2 Department of Crop Sciences, Research Institute of Organic Agriculture (FiBL), Ackerstrasse 113, 5070 Frick, Switzerland 3 INRA, Centre de Recherche PACA, UR Ecodeveloppement, 84914 Avignon, France 4 Department of Plant Protection Biology—Unit of Integrated Plant Protection, Swedish University of Agricultural Science, P.O. Box 102, SE-230 53 Alnarp, Sweden 5 Department of Plant and Environmental Sciences, University of Copenhagen (UCPH), Thorvaldsensvej 40, DK-1871 Frederiksberg C, Denmark * Correspondence: [email protected] Received: 31 May 2019; Accepted: 6 August 2019; Published: 11 August 2019 Abstract: Functional biodiversity is of fundamental importance for pest control. Many natural enemies rely on floral resources to complete their life cycle. Farmers need to ensure the availability of suitable and sufficient floral biodiversity. This review summarizes 66 studies on the management of floral biodiversity in apple orchards, published since 1986. Approaches followed different degrees of intervention: short-term practices (mowing regime and weed maintenance, cover crops), establishment of durable ecological infrastructures (perennial flower strips, hedgerows) and re-design of the crop system (intercropping, agroforestry). Although short-term practices did not always target the nutrition of natural enemies by flowering plants, living conditions for them (alternative prey, provision of habitat) were often improved. Perennial flower strips reliably enhanced natural enemies and techniques for their introduction continuously developed. -

1 Appendix 3. Gulf Islands Taxonomy Report
Appendix 3. Gulf Islands Taxonomy Report Class Order Family Genus Species Arachnida Araneae Agelenidae Agelenopsis Agelenopsis utahana Eratigena Eratigena agrestis Amaurobiidae Callobius Callobius pictus Callobius severus Antrodiaetidae Antrodiaetus Antrodiaetus pacificus Anyphaenidae Anyphaena Anyphaena aperta Anyphaena pacifica Araneidae Araneus Araneus diadematus Clubionidae Clubiona Clubiona lutescens Clubiona pacifica Clubiona pallidula Cybaeidae Cybaeus Cybaeus reticulatus Cybaeus signifer Cybaeus tetricus Dictynidae Emblyna Emblyna peragrata Gnaphosidae Sergiolus Sergiolus columbianus Zelotes Zelotes fratris Linyphiidae Agyneta Agyneta darrelli Agyneta fillmorana Agyneta protrudens Bathyphantes Bathyphantes brevipes Bathyphantes keeni 1 Centromerita Centromerita bicolor Ceratinops Ceratinops latus Entelecara Entelecara acuminata Erigone Erigone aletris Erigone arctica Erigone cristatopalpus Frederickus Frederickus coylei Grammonota Grammonota kincaidi Linyphantes Linyphantes nehalem Linyphantes nigrescens Linyphantes pacificus Linyphantes pualla Linyphantes victoria Mermessus Mermessus trilobatus Microlinyphia Microlinyphia dana Neriene Neriene digna Neriene litigiosa Oedothorax Oedothorax alascensis Pityohyphantes Pityohyphantes alticeps Pocadicnemis Pocadicnemis pumila Poeciloneta Poeciloneta fructuosa Saaristoa Saaristoa sammamish Scotinotylus Scotinotylus sp. 5GAB Semljicola Semljicola sp. 1GAB Sisicottus Spirembolus Spirembolus abnormis Spirembolus mundus Tachygyna Tachygyna ursina Tachygyna vancouverana Tapinocyba Tapinocyba