Patterns of Plant Composition in Fragments of Globally Imperiled
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

Sistema De Clasificación Artificial De Las Magnoliatas Sinántropas De Cuba
Sistema de clasificación artificial de las magnoliatas sinántropas de Cuba. Pedro Pablo Herrera Oliver Tesis doctoral de la Univerisdad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2007 Sistema de clasificación artificial de las magnoliatas sinántropas de Cuba. Pedro Pablo Herrera Oliver PROGRAMA DE DOCTORADO COOPERADO DESARROLLO SOSTENIBLE: MANEJOS FORESTAL Y TURÍSTICO UNIVERSIDAD DE ALICANTE, ESPAÑA UNIVERSIDAD DE PINAR DEL RÍO, CUBA TESIS EN OPCIÓN AL GRADO CIENTÍFICO DE DOCTOR EN CIENCIAS SISTEMA DE CLASIFICACIÓN ARTIFICIAL DE LAS MAGNOLIATAS SINÁNTROPAS DE CUBA Pedro- Pabfc He.r retira Qltver CUBA 2006 Tesis doctoral de la Univerisdad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2007 Sistema de clasificación artificial de las magnoliatas sinántropas de Cuba. Pedro Pablo Herrera Oliver PROGRAMA DE DOCTORADO COOPERADO DESARROLLO SOSTENIBLE: MANEJOS FORESTAL Y TURÍSTICO UNIVERSIDAD DE ALICANTE, ESPAÑA Y UNIVERSIDAD DE PINAR DEL RÍO, CUBA TESIS EN OPCIÓN AL GRADO CIENTÍFICO DE DOCTOR EN CIENCIAS SISTEMA DE CLASIFICACIÓN ARTIFICIAL DE LAS MAGNOLIATAS SINÁNTROPAS DE CUBA ASPIRANTE: Lie. Pedro Pablo Herrera Oliver Investigador Auxiliar Centro Nacional de Biodiversidad Instituto de Ecología y Sistemática Ministerio de Ciencias, Tecnología y Medio Ambiente DIRECTORES: CUBA Dra. Nancy Esther Ricardo Ñapóles Investigador Titular Centro Nacional de Biodiversidad Instituto de Ecología y Sistemática Ministerio de Ciencias, Tecnología y Medio Ambiente ESPAÑA Dr. Andreu Bonet Jornet Piiofesjar Titular Departamento de EGdfegfe Universidad! dte Mearte CUBA 2006 Tesis doctoral de la Univerisdad de Alicante. Tesi doctoral de la Universitat d'Alacant. 2007 Sistema de clasificación artificial de las magnoliatas sinántropas de Cuba. Pedro Pablo Herrera Oliver I. INTRODUCCIÓN 1 II. ANTECEDENTES 6 2.1 Historia de los esquemas de clasificación de las especies sinántropas (1903-2005) 6 2.2 Historia del conocimiento de las plantas sinantrópicas en Cuba 14 III. -

Cocoa Beach Maritime Hammock Preserve Management Plan
MANAGEMENT PLAN Cocoa Beach’s Maritime Hammock Preserve City of Cocoa Beach, Florida Florida Communities Trust Project No. 03 – 035 –FF3 Adopted March 18, 2004 TABLE OF CONTENTS SECTION PAGE I. Introduction ……………………………………………………………. 1 II. Purpose …………………………………………………………….……. 2 a. Future Uses ………….………………………………….…….…… 2 b. Management Objectives ………………………………………….... 2 c. Major Comprehensive Plan Directives ………………………..….... 2 III. Site Development and Improvement ………………………………… 3 a. Existing Physical Improvements ……….…………………………. 3 b. Proposed Physical Improvements…………………………………… 3 c. Wetland Buffer ………...………….………………………………… 4 d. Acknowledgment Sign …………………………………..………… 4 e. Parking ………………………….………………………………… 5 f. Stormwater Facilities …………….………………………………… 5 g. Hazard Mitigation ………………………………………………… 5 h. Permits ………………………….………………………………… 5 i. Easements, Concessions, and Leases …………………………..… 5 IV. Natural Resources ……………………………………………..……… 6 a. Natural Communities ………………………..……………………. 6 b. Listed Animal Species ………………………….…………….……. 7 c. Listed Plant Species …………………………..…………………... 8 d. Inventory of the Natural Communities ………………..………….... 10 e. Water Quality …………..………………………….…..…………... 10 f. Unique Geological Features ………………………………………. 10 g. Trail Network ………………………………….…..………..……... 10 h. Greenways ………………………………….…..……………..……. 11 i Adopted March 18, 2004 V. Resources Enhancement …………………………..…………………… 11 a. Upland Restoration ………………………..………………………. 11 b. Wetland Restoration ………………………….…………….………. 13 c. Invasive Exotic Plants …………………………..…………………... 13 d. Feral -

New Sesquiterpene Lactones from the Genera Calea and Berlandiera (Asteraceae) and the Fragmentation Reactions of 1,3-Dihydroxyeudesmanolide Derivatives
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1983 New Sesquiterpene Lactones From the Genera Calea and Berlandiera (Asteraceae) and the Fragmentation Reactions of 1,3-Dihydroxyeudesmanolide Derivatives. Ihl Young Lee Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Lee, Ihl Young, "New Sesquiterpene Lactones From the Genera Calea and Berlandiera (Asteraceae) and the Fragmentation Reactions of 1,3-Dihydroxyeudesmanolide Derivatives." (1983). LSU Historical Dissertations and Theses. 3895. https://digitalcommons.lsu.edu/gradschool_disstheses/3895 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. INFORMATION TO USERS This reproduction was made from a copy of a document sent to us for microfilming. While the most advanced technology has been used to photograph and reproduce this document, the quality of the reproduction is heavily dependent upon the quality of the material submitted. The following explanation of techniques is provided to help clarify markings or notations which may appear on this reproduction. 1.The sign or “target” for pages apparently lacking from the document photographed is “Missing Page(s)”. If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting through an image and duplicating adjacent pages to assure complete continuity. 2. -

FINAL REPORT PSRA Vegetation Monitoring 2005-2006 PC P502173
Rare Plants and Their Locations at Picayune Strand Restoration Area: Task 4a FINAL REPORT PSRA Vegetation Monitoring 2005-2006 PC P502173 Steven W. Woodmansee and Michael J. Barry [email protected] December 20, 2006 Submitted by The Institute for Regional Conservation 22601 S.W. 152 Avenue, Miami, Florida 33170 George D. Gann, Executive Director Submitted to Mike Duever, Ph.D. Senior Environmental Scientist South Florida Water Management District Fort Myers Service Center 2301 McGregor Blvd. Fort Myers, Florida 33901 Table of Contents Introduction 03 Methods 03 Results and Discussion 05 Acknowledgements 38 Citations 39 Tables: Table 1: Rare plants recorded in the vicinity of the Vegetation Monitoring Transects 05 Table 2: The Vascular Plants of Picayune Strand State Forest 24 Figures: Figure 1: Picayune Strand Restoration Area 04 Figure 2: PSRA Rare Plants: Florida Panther NWR East 13 Figure 3: PSRA Rare Plants: Florida Panther NWR West 14 Figure 4: PSRA Rare Plants: PSSF Northeast 15 Figure 5: PSRA Rare Plants: PSSF Northwest 16 Figure 6: PSRA Rare Plants: FSPSP West 17 Figure 7: PSRA Rare Plants: PSSF Southeast 18 Figure 8: PSRA Rare Plants: PSSF Southwest 19 Figure 9: PSRA Rare Plants: FSPSP East 20 Figure 10: PSRA Rare Plants: TTINWR 21 Cover Photo: Bulbous adder’s tongue (Ophioglossum crotalophoroides), a species newly recorded for Collier County, and ranked as Critically Imperiled in South Florida by The Institute for Regional Conservation taken by the primary author. 2 Introduction The South Florida Water Management District (SFWMD) plans on restoring the hydrology at Picayune Strand Restoration Area (PSRA) see Figure 1. -

Galactia Smallii) and Candidate Sand Flax (Linum Arenicola) Dade County Florida, Adjacent to SOCSOUTH Headquarters Homestead, Florida
Plant Survey for the Endangered Small’s Milkpea (Galactia smallii) and Candidate Sand Flax (Linum arenicola) Dade County Florida, Adjacent to SOCSOUTH Headquarters Homestead, Florida PO 952722 Craig van der Heiden and James Johnson October 30, 2013 Small’s milkpea Sand flax Submitted by: The Institute for Regional Conservation 100 E. Linton Blvd, Suite 302B Delray Beach, FL 33483 Craig van der Heiden PhD, CEO Submitted to: CH2M HILL Northpark 400 1000 Abernathy Road, Suite 1600 Atlanta, GA 30328 2 Table of Contents Introduction ......................................................................................................................... 3 Methods............................................................................................................................... 3 Results ................................................................................................................................. 4 Survey area 1................................................................................................................... 5 Survey area 2................................................................................................................... 6 Survey area 3................................................................................................................... 7 Survey area 4................................................................................................................... 9 Survey area 5................................................................................................................ -
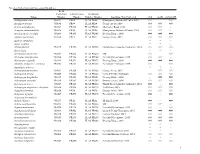
Species Collection and Accession Numbers
Species Collection and Accession Numbers Field Collection Identification Herbarium Taxon Number Number Voucher Number Location; Year Collected rbcL matK psbA-trnH Abildgaardia ovata 950034 PR60 FLAS PR60 Montgomery Botanical Center; 2014 ### - ### Abrus precatorius 950080 PR84 FLAS PR84 George Avery; 2014 ### ### ### Acacia auriculformis 950414 PR398 FLAS PR398 Old Culter Road; 2015 ### ### - Acalypha chamaedrifolia 950036 PR50 FLAS PR50 Montgomery Botanical Center; 2014 ### ### - Aeschynomene viscidula 950209 PR145 FLAS PR145 Deering Estate; 2014 ### ### ### Agalinis fasciculata 950052 PR71 FLAS PR71 George Avery; 2014 ### ### ### Agalinis obtusifolia * * - Agave sisalana * * - Albizia lebbeck 950225 PR198 FLAS PR198 Florida Keys Aquaduct Authority; 2014 ### ### ### Aletris lutea - * - Alternathera flavescens 950244 PR252 FLAS PR252 SM ### - ### Alvaradoa amorphoides 950260 PR244 FLAS PR244 Fairchild Greenhouse; 2015 ### ### - Alysicarpus vaginalis 950169 PR173 FLAS PR173 Deering Estate; 2014 ### ### ### Amorpha herbacea v. crenulata 950258 PR238 FLAS PR238 Fairchild Greenhouse; 2015 ### - ### Ampelopsis arborea * * - Andropogon glomeratus 950063 PR126 FLAS PR126 George Avery; 2014 - ### ### Andropogon gyrans 950445 PR466 FLAS PR466 Larry & Penny Thompson ### ### ### Andropogon longiberbis 950187 PR151 FLAS PR151 Deering Estate; 2014 - ### ### Andropogon ternarius 950189 PR163 FLAS PR163 Deering Estate; 2014 ### ### ### Andropogon tracyi 950227 PR160 FLAS PR160 Florida Keys Aquaduct Authority; 2014 ### - ### Andropogon virginicus v. decipiens -

In the Flora of South Florida
. PlQt!JRe?\ATE Report T-558 Endemic Taxa,-inthe Flora of South Florida*' NATIONAL Y Everglades National Park, South Florida Research Center, P.O. Box 279, Homestead, Florida 33030 I, ,. ,. ,#< Endemic Taxa in the Flora of South Florida " - Report T-558 George N. Avery and Lloyd L. Loope . U.S. National Park Service ' South Florida Research Center Everglades National Park Homestead, Florida 33030 July 1980 . Avery, George N. and Lloyd L. Loope. 1980. ~ndemicTaxa in the Flora of South Florida. South Florida Research Center Report T-558. 39 pp. Endemic Taxa in the Flora of South Florida TABLE OF CONTENTS Page INTRODUCTION . 1 LITERATURE ON SOUTH FLORIDA ENDEMICS . METHODS . rr , ANNOTATED LIST OF THE ENDEMIC SOUTH FLORIDA FLORA . DISCUSSION. I . \ '& ACKNOWLEDGEMENTS ........................ LITERATURE CITED . 18 Table 1. Habitat and conservation status of endemic plant taxa of.SoutH Florida . .. 6. Table 2. Number of endemics found in selected vegetation categories . APPENDIX I - Annotated ,version of Robertson's (1955) list of South Florida endemics, showing .diff erences from our list . : Endemic Taxa in the Flora of South Florida George N. Avery and kloyd L. Loope , INTRODUCTION The island-like tropical area of South Florida possesses a very remarkable flora by North American standards, with a high percentage of species having tropical affinities and with fairly high local endemism. Hundreds of plant species known from the United States are found only in Florida south of Lake Okeechobee. Many of these species occur on various Caribbean islands and elsewhere in the Neotropics. This report treats those taxa endemic to South Florida, occurring in peninsular Florida southbf Lake Okeechobee and/or on the Florida Keys, and found nowhere else. -

Pineland Golden Trumpet (Angadenia Berteroi)
Pineland golden trumpet (Angadenia berteroi) For definitions of botanical terms, visit en.wikipedia.org/wiki/Glossary_of_botanical_terms. Pineland golden trumpet is an erect or vine-like wildflower found in pinelands, dry hammocks and marl prairies of Monroe County and the Keys. It is a state-listed threatened species. Its creamy yellow flowers typically bloom in spring and summer, but may bloom throughout the year. They attract a variety of insects, especially moths, which are its primary pollinator. The plant is a larval host for the Oleander and Polka dot wasp Photo by Scott Zona (CC BY-NC 2.0) moths. The flower is trumpet-shaped with a broad, five- lobed corolla. The lobes overlap, giving the flower a pinwheel-like appearance. Red lines mark the interior of the floral tube. Five stamens are fused to the tube and enclose the stigma to increase the likelihood of pollination. Leaves are linear to oblong, glabrous and leathery with entire, slightly revolute margins. They are oppositely arranged. Stems are smooth and produce a milky sap when broken. The fruit is a cylindrical follicle and is born in pairs. Seeds are flat, brown and bear a silky pappus that catches the wind to aid in dispersal. Family: Apocynaceae (Dogbane family) Native range: Miami-Dade County, the Keys To see where natural populations of Pineland golden trumpet have been vouchered, visit www.florida.plantatlas.usf.edu. Hardiness: Zones 10B–11 Lifespan: Perennial Soil: Dry to moist, well-drained calcareous or sandy soils Exposure: Full sun to partial shade Growth habit: 6–24” tall, or up to 3’ long Propagation: Seeds Garden tips: Pineland golden trumpet is suitable for use in a wildflower garden. -

PLANT SCIENCE Bulletin Fall 2013 Volume 59 Number 3
PLANT SCIENCE Bulletin Fall 2013 Volume 59 Number 3 Botany in Action - in New Orleans! In This Issue.............. More BSA awards announced at PlantingScience mentors Botany 2013!.....p. 146 Botany 2013.....p. 80 make a difference.....p. 90 From the Editor PLANT SCIENCE BULLETIN The good news these days is about resources. There is so much information readily available on the internet Editorial Committee that one hardly needs to leave the office to work on a Volume 59 literature review or gather information for a lecture. The first step—Google it! The bad news these days is Elizabeth Schussler about resources. There is so much information readily (2013) available on the internet that one could spend hours Department of Ecology & sorting through possible sites to find the information Evolutionary Biology you want. What we need is a resource that has done University of Tennessee the dirty work of searching what is available and evalu- Knoxville, TN 37996-1610 ating its usefulness. That resource has been provided [email protected] for botanical and lichenological systematic research by Morgan Gostel, Manuela Dal-Forno, and Andrea Weeks in this issue. This is also a great resource to use for teaching images. Christopher Martine (2014) In our other feature article, Melanie Link-Pérez and Department of Biology Elizabeth Schussler demonstrate that resources, by Bucknell University themselves, are not enough to support grade-school Lewisburg, PA 17837 teachers in their efforts to introduce plant science to [email protected] students. At this age the kids love plants and so do the teachers, and the teachers are anxious to find and use resources to help them incorporate plants into the curriculum. -

1 DEPARTMENT of the INTERIOR Fish and Wildlife
This document is scheduled to be published in the Federal Register on 08/04/2016 and available online at http://federalregister.gov/a/2016-17322, and on FDsys.gov DEPARTMENT OF THE INTERIOR Fish and Wildlife Service 50 CFR Part 17 [Docket No. FWS–R9–ES–2008–0063; 92300-1113-0000-9B] RIN 1018–AU62 Endangered and Threatened Wildlife and Plants; Amending the Formats of the Lists of Endangered and Threatened Wildlife and Plants AGENCY: Fish and Wildlife Service, Interior. ACTION: Final rule. SUMMARY: We, the U.S. Fish and Wildlife Service, amend the format of the Lists of Endangered and Threatened Wildlife and Plants (Lists) to reflect current practices and standards that will make the regulations and Lists easier to understand. The Lists, in the new format, are included in their entirety and have been updated to correct identified errors. 1 DATES: This rule is effective [INSERT DATE OF PUBLICATION IN THE FEDERAL REGISTER]. FOR FURTHER INFORMATION CONTACT: Don Morgan, Ecological Services Program, U.S. Fish and Wildlife Service, 5275 Leesburg Pike, Falls Church, VA, 22041; telephone 703– 358–2171. If you use a telecommunications device for the deaf (TDD), call the Federal Information Relay Service (FIRS) at 800–877–8339. SUPPLEMENTARY INFORMATION: Background The Lists of Endangered and Threatened Wildlife and Plants (Lists), found in title 50 of the Code of Federal Regulations (CFR) at 50 CFR 17.11 for wildlife and 50 CFR 17.12 for plants, contain the names of endangered species and threatened species officially listed pursuant to the Endangered Species Act of 1973, as amended (16 U.S.C. -

Appendix A. Plant Species Known to Occur at Canaveral National Seashore
National Park Service U.S. Department of the Interior Natural Resource Stewardship and Science Vegetation Community Monitoring at Canaveral National Seashore, 2009 Natural Resource Data Series NPS/SECN/NRDS—2012/256 ON THE COVER Pitted stripeseed (Piriqueta cistoides ssp. caroliniana) Photograph by Sarah L. Corbett. Vegetation Community Monitoring at Canaveral National Seashore, 2009 Natural Resource Report NPS/SECN/NRDS—2012/256 Michael W. Byrne and Sarah L. Corbett USDI National Park Service Southeast Coast Inventory and Monitoring Network Cumberland Island National Seashore 101 Wheeler Street Saint Marys, Georgia, 31558 and Joseph C. DeVivo USDI National Park Service Southeast Coast Inventory and Monitoring Network University of Georgia 160 Phoenix Road, Phillips Lab Athens, Georgia, 30605 March 2012 U.S. Department of the Interior National Park Service Natural Resource Stewardship and Science Fort Collins, Colorado The National Park Service, Natural Resource Stewardship and Science office in Fort Collins, Colorado publishes a range of reports that address natural resource topics of interest and applicability to a broad audience in the National Park Service and others in natural resource management, including scientists, conservation and environmental constituencies, and the public. The Natural Resource Data Series is intended for the timely release of basic data sets and data summaries. Care has been taken to assure accuracy of raw data values, but a thorough analysis and interpretation of the data has not been completed. Consequently, the initial analyses of data in this report are provisional and subject to change. All manuscripts in the series receive the appropriate level of peer review to ensure that the information is scientifically credible, technically accurate, appropriately written for the intended audience, and designed and published in a professional manner.