Download This PDF File
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
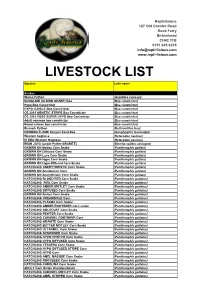
Livestock List
Repti-lisious 187 Old Chester Road Rock Ferry Birkenhead CH42 3TB 0151 645 6235 [email protected] www.repti-lisious.com LIVESTOCK LIST Species Latin name Snakes Column1 Woma Python (Aspidites ramsayi) SUNGLOW (ALBINO SHARP) Boa (Boa constrictor) Hypo Boa Constrictor (Boa constrictor) HYPO JUNGLE Boa Constrictor (Boa constrictor) CB 2015 GENETIC STRIPE Boa Constrictor (Boa constrictor) CB 2015 POSS SUPER HYPO Boa Constrictor (Boa constrictor) Adult common boa constrictor (Boa constrictor) Blood salmon boa constrictor (Boa constrictor) Bismark Python (Bothrochilus boa) DODOMA FLAME Kenyan Sand Boa (Gonglyophis loveredgei) Western hognose (Heterodon nasicus) HI RED Western Hognose (Heterodon nasicus) IRIAN JAYA Carpet Py(het GRANITE) (Morelia spilota variagata) GROWN ON Motley Corn Snake (Pantherophis guttata) GROWN ON Diffused Corn Snake (Pantherophis guttata) GROWN ON Lava Corn Snake (Pantherophis guttata) GROWN ON Hypo Corn Snake (Pantherophis guttata) GROWN ON Hypo Diffused Corn Snake (Pantherophis guttata) HATCHLING ANERYTHRISTIC Corn Snake (Pantherophis guttata) GROWN ON Amelanistic Corn (Pantherophis guttata) GROWN ON Anerythristic Corn Snake (Pantherophis guttata) HATCHLING BLOOD RED Corn Snake (Pantherophis guttatus) HATCHLING FIRE Corn Snake (Pantherophis guttatus) HATCHLING AMBER MOTLEY Corn Snake (Pantherophis guttatus) HATCHLING DIFFUSED Corn Snake (Pantherophis guttatus) GROWN ON Amber Corn Snake (Pantherophis guttatus) HATCHLING CREAMSICLE Corn (Pantherophis guttatus) HATCHLING PLASMA Corn Snake (Pantherophis guttatus) -

A. Kumbang Dewasa
Keanekaragaman KUMBANG STAG (Coleoptera: Lucanidae) di Pulau Jawa Woro Anggraitoningsih Noerdjito i Dilarang mereproduksi atau memperbanyak seluruh atau sebagian dari buku ini dalam bentuk atau cara apa pun tanpa izin tertulis dari penerbit. © Hak cipta dilindungi oleh Undang-Undang No. 28 Tahun 2014 All Rights Reserved ii Keanekaragaman KUMBANG STAG (Coleoptera: Lucanidae) di Pulau Jawa Woro Anggraitoningsih Noerdjito LIPI Press iii © 2016 Lembaga Ilmu Pengetahuan Indonesia Pusat Penelitian Biologi Katalog dalam terbitan Keanekaragaman Kumbang Stag (Coleoptera: Lucanidae) di Pulau Jawa/Woro Anggraitoningsih Noerdjito – Jakarta: LIPI Press, 2016. xxiv+148; 14,8 x 21 cm. ISBN 978-979-799-856-1 1. Keanekaragaman 2. Kumbang stag 595.764 9 Copy editor : M. Sidik Nugraha dan Sonny Heru Kusuma Proofreader : Sarwendah Puspita Dewi dan Fadly Suhendra Penata Isi : Erna Rumbiati, Prapti Sasiwi, dan Rahma Hilma Taslima Desainer sampul : Rusli Fazi Cetakan pertama : November 2016 Diterbitkan oleh: LIPI Press, anggota Ikapi Jln. Gondangdia Lama 39, Menteng, Jakarta 10350 Telp.: (021) 314 0228, 314 6942. Faks.: (021) 314 4591 E-mail: [email protected] Website: lipipress.lipi.go.id LIPI Press @lipi_press iv DAFTAR ISI DAFTAR GAMBAR .................................................................................vii DAFTAR TABEL ......................................................................................xv PENGANTAR PENERBIT.....................................................................xvii KATA PENGANTAR ...............................................................................xix -

Sebastian, Zimmer Bankdaten
International-Insect Groß-Umstädterstr.38 64823 Groß-Umstadt +4915212129805 Geschäftsführer: Sebastian, Zimmer Bankdaten: Sebastian Zimmer Volksbank Odenwald IBan:DE94508635130005322456 BIC:GENODE51MIC Species Name Order SNAILS NO.1001 Archachatina marginata ovum 16cm +/- Giant African Snail Archachatina m. ovum 15 cm Giant African Snail NO.1002 Archachatina marginata ovum albino Giant African Snail Albino coming soon NO.1003 Archachatina marginata ovum ( albina ) Yellow shell NO.1004 Achatina achatina Tiger Snail NO.1005 Edentulina Obesum Predator snail coming soon NO.1006 Archachatina marginata Sulturalis Pink Lip Snail NO.1007 achatina achatina albino Giant African Snail Albino NO.1008 Archachatina species Pink IKOM coming soon NO.1009 Limicolaria flammea With Shell coming soon Limicularia flammea pink shell coming soon Limicolaria Atheopis Fire flame coming soon Limicolaria White shell marbel coming soon Limicolaria Giant Mixed color coming soon Limicolaria flammea stripe shell coming soon NO.1011 veronicella sp Pan cake Slug Thailand coming soon NO.1012 Archachatina pulaerty Sylver Body coming soon NO.1013 Archachatina Knorry Red tail snail coming soon NO.1014 Archachatina marginata marginata Cameron Border Snails Archachatina papyracea Wild colore coming soon Archachatina papyracea Albino shell Pseudoachatina sp. Very nice very rare coming soon Archachatina Ventricosa Purpel mouth Archachatina sp. marbel shell coming soon Archachatina cf papyracea New and good to Keep coming soon Achatina balteata Stripe body Snail Import Coming -

Official Lists and Indexes of Names and Works in Zoology
OFFICIAL LISTS AND INDEXES OF NAMES AND WORKS IN ZOOLOGY Supplement 1986-2000 Edited by J. D. D. SMITH Copyright International Trust for Zoological Nomenclature 2001 ISBN 0 85301 007 2 Published by The International Trust for Zoological Nomenclature c/o The Natural History Museum Cromwell Road London SW7 5BD U.K. on behalf of lICZtN] The International Commission on Zoological Nomenclature 2001 STATUS OF ENTRIES ON OFFICIAL LISTS AND INDEXES OFFICIAL LISTS The status of names, nomenclatural acts and works entered in an Official List is regulated by Article 80.6 of the International Code of Zoological Nomenclature. All names on Official Lists are available and they may be used as valid, subject to the provisions of the Code and to any conditions recorded in the relevant entries on the Official List or in the rulings recorded in the Opinions or Directions which relate to those entries. However, if a name on an Official List is given a different status by an adopted Part of the List of Available Names in Zoology the status in the latter is to be taken as correct (Article 80.8). A name or nomenclatural act occurring in a work entered in the Official List of Works Approved as Available for Zoological Nomenclature is subject to the provisions of the Code, and to any limitations which may have been imposed by the Commission on the use of that work in zoological nomenclature. OFFICIAL INDEXES The status of names, nomenclatural acts and works entered in an Official Index is regulated by Article 80.7 of the Code. -

Artikel-Preisliste Käfer
[email protected] Artikel-Preisliste Tel.: +49 5043 98 99 747 Fax: +49 5043 98 99 749 Weitere Informationen zu unseren Produkten sowie aktuelle Angebote finden Sie in unserem Shop: www.thepetfactory.de Art.-Nr. Artikelbezeichnung Ihr Preis MwSt.-Satz * Käfer - Imagines AEGP Aegus parallelus - Pärchen 39,95 € 1 ANTSXMA Anthia sexmaculata - Stückpreis 17,50 € 1 APHBELIM Stephanorrhina bella - Stückpreis 14,95 € 1 CETSPK1 Pseudinca camerunensis - Stückpreis 14,95 € 1 CETSPK2 Plaesiorrhinella watkinsiana - Stückpreis 15,95 € 1 CHACAMI Chlorocala africana camerunica - Stückpreis 7,95 € 1 CHASUB Chlorocala africana subsuturalis - Stückpreis 14,50 € 1 CHAT Chalcosoma atlas - Pärchen 34,95 € 1 CHATXL Chalcosoma atlas XL - Pärchen 49,95 € 1 CHCA Chalcosoma caucasus - Pärchen 39,95 € 1 CHCAX Chalcosoma caucasus XXL - Pärchen 69,95 € 1 CHCON Chlorocala conjux - Stückpreis 17,50 € 1 COLORI Coelorrhina loricata Imago - Stückpreis 19,95 € 1 DEPCAN Depilicus cantori 49,00 € 1 DIMIIM Dicronorhina micans - Pärchen 39,95 € 1 DIPGAGA Diplognatha cf. gagates 14,50 € 1 DIPSILI Diplognatha silicea - Stückpreis 14,50 € 1 DOAL Dorcus alcides - Pärchen 24,95 € 1 DOBU Dorcus bucephalus - Pärchen 42,00 € 1 DOREI Dorcus reichei - Pärchen 29,95 € 1 DOTA Dorcus taurus - Pärchen 39,50 € 1 DOTS Dorcus titanus SUMATRA - Pärchen 26,95 € 1 DOTSUL Dorcus titanus titanus SULAWESI - Pärchen 26,95 € 1 GYMMAR Gymnetis marmorea 14,95 € 1 HEBU Hexarthrius buqueti - Pärchen 32,50 € 1 HEPA Hexarthrius parryi - Pärchen 39,50 € 1 HOGL Homoderus gladiator - Pärchen 39,50 -

Diplomarbeit
DIPLOMARBEIT Titel der Diplomarbeit „Successional patterns of necrophilous beetles on domestic pig carcasses in urban and sylvan areas during spring and summer – a comparative study between four study sites in and around Vienna“ Verfasser Anatole Johannes Patzak angestrebter akademischer Grad Magister der Naturwissenschaften (Mag.rer.nat.) Wien, 2011 Studienkennzahl lt. A 444 Studienblatt: Studienrichtung lt. Ökologie Studienblatt: Betreuer: O. Univ.-Prof. Dr. Hannes Paulus Ao. Univ.-Prof. Dr. Christian Reiter “I have always looked upon decay as being just as wonderful and rich an expression of life as growth.” Henry Miller 1 INDEX 1. ABSTRACT ...................................................................................................................... 5 Zusammenfassung ......................................................................................................... 7 2. INTRODUCTION ............................................................................................................... 9 2.1 Forensic entomology ................................................................................................ 9 2.2 Carrion ecology ...................................................................................................... 10 2.2.1 Characteristics of a carcass ............................................................................... 11 2.2.2 Carrion as a habitat ........................................................................................... 11 2.2.3 Stages of decomposition .................................................................................. -

Conservation Milestones of the Critically Endangered Philippine Crocodile (Crocodylus Mindorensis Schmidt 1935)
SYLVATROP Editorial Staff Bighani M. Manipula, PhD Melanie N. Ojeda Editor-in-Chief Editorial Assistant Adreana Santos-Remo Marilou C. Villones Managing Editor Circulation Officer/Proofreader Liberty E. Asis Eduardo M. Tolentino Adreana Santos-Remo Catalina DM. Aldemita Editors Gino S. Laforteza Circulation Assistants Maria Lourdes Q. Moreno, PhD Guest Editor Liberato A. Bacod Printing Supervisor January - June 2018 Vol. 28 No. 1 SYLVATROP, The Technical Journal of Philippine Ecosystems and Natural Resources is published by the Department of Environment and Natural Resources (DENR) through the Ecosystems Research and Development Bureau (ERDB), College, Laguna. Sylvatrop is listed in Clarivate Analytics (formerly Thomson Reuters) Master Journal List. Subscription rates: Php75 for single issue copy (local); Php150 for combined issues and US$15 per single issue copy (foreign) excluding airmail cost; US$30 for combined issues. Re-entered as Second Class Mail CY 2018 at the College, Laguna Post Office on 23 March 2018. Permit No. 2018-21. Address checks to Ecosystems Research and Development Bureau c/o ERDB Circulation. For contributions or inquiries, address it to The Editor-in-Chief at the following address: SYLVATROP, The Technical Journal of Philippine Ecosystems and Natural Resources Ecosystems Research and Development Bureau, DENR Tel. No. (049) 557-1758 Fax: (049) 536-2850 E-mail address: [email protected]; [email protected] Website: sylvatrop.denr.gov.ph Cover Photo: The cover photo shows the species [Buceros hydrocorax, Gallirallus calayanensis, Prionailurus bengalensis heaneyi, Pithecophaga jefferyi, Pteropus pumilus, Otus megalotis, Crocodylus mindorensis (top to bottom)] included in the articles focusing on species conservation. Photo Credit: R. Sta. -

Pola Populasi Kumbang Lucanid (Coleoptera:Lucanidae) Pada Berbagai Tipe Habitat Di Hutan Gunung Salak, Jawa Barat
POLA POPULASI KUMBANG LUCANID (COLEOPTERA:LUCANIDAE) PADA BERBAGAI TIPE HABITAT DI HUTAN GUNUNG SALAK, JAWA BARAT Roni Koneri 1) 1)Program Studi Biologi, FMIPA, Universitas Sam Ratulangi Jalan Kampus Unsrat, Manado 95115 e-mail: [email protected] ABSTRAK Penelitian ini bertujuan untuk mengkaji pola populasi kumbang lucanid di hutan Gunung Salak. Data populasi kumbang lucanid diperoleh dengan menggunakan perangkap lampu (light trap). Penelitian dilaksanakan di kawasan hutan Gunung Salak dan berlangsung dari bulan Mei 2004 sampai Desember 2005. Hasil penelitian menunjukkan bahwa kelimpahan populasi di alam mencapai puncaknya pada bulan April dan Mei. Spesies yang selalu muncul setiap bulan selama pengamatan adalah Cyclomatus canaliculatus, Prosopocoilus astocoides dan Hexarthrius buqueti. Kata kunci: Gunung Salak, kumbang lucanid, populasi. POPULATION PATTERN OF LUCANID BEETLE IN DIFFERENT HABITAT TYPE AT SALAK MOUNTAIN, WEST JAVA ABSRACT The objectives of the research were to study temporal dynamic pattern of Lucanid beetle in different habitat type. Research was conducted between May 2004 and December 2005. Beetles were surveyed by using light traps in three different forest types. Results have indicated that temporal dynamic pattern of Lucanid beetle was affected by forest disturbance level. Highest number of Lucanid beetles were recorded in April dan May. Species which always emerge each month during perception is Cyclomatus canaliculatus, Prosopocoilus astocoides and Hexarthrius buqueti. Keywords: Salak mountain, lucanids beetle, population. PENDAHULUAN fluktuasi serangga telah dilaporkan, seperti: pengaruh perangkap cahaya terhadap Populasi kumbang di alam fluktuasi kumbang permukaan tanah (ground dipengaruhi oleh dinamika lingkungannya. beetle) selama sepuluh tahun (Yahiro & Dinamika yang terjadi berupa perubahan Yano, 1997), hubungan antara iklim dengan fenologi tumbuhan hutan, kondisi fisik, fluktuasi kupu-kupu (Woong, 2003), iklim dari waktu ke waktu mempengaruhi hubungan antara teknik pengambilan sampel reproduksi, pertumbuhan dan mortalitasnya. -

Kuota Penangkapan Jenis Satwa Dan Pengambilan Jenis Tumbuhan Appendiks Ii Cites Periode 2016
Lampiran Surat No. : SK. 283/KSDAE-SET/2015 Tanggal : 11 Desember 2015 KUOTA PENANGKAPAN JENIS SATWA DAN PENGAMBILAN JENIS TUMBUHAN APPENDIKS II CITES PERIODE 2016 Jatah Lokasi No. Nama Jenis Keterangan Tangkap Export Tangkap/Ambil MAMALIA 1 Pteropus vampyrus 250 225 Total Common Flying-fox Sumut 200 Kalong Kapauk 50 Jateng REPTILIA ULAR / SNAKES 1 Liasis (Apodora) papuanus 300 270 Total Papuan Olive Python / 200 Papua Barat Hidup (Pet) Papua Ular Sanca Irian 100 2 Candoia aspera 1.200 1.080 Total Papuan Ground Boa / 800 Papua Barat Hidup (Pet) Ular Boa Tanah 400 Papua 3 Candoia carinata 600 540 Total Papuan Tree Boa / Hidup (Pet) Ular Boa Pohon 350 Papua Barat 250 Papua 4 Candoia paulsoni 200 180 Total Halmahera Ball Boa / 200 Maluku Utara Hidup (Pet) Ular Boa Halmahera (Halmahera) 5 Bothrochilus 500 450 Total (Leiopython) albertisi 300 Papua Barat Hidup (Pet) White-lipped / D'Albert's 200 Papua Ular Sanca Bibir Putih 6 Total Simalia (Morelia) amethistina 150 135 Scrub / Amesthistine Python 50 Papua Barat Hidup (Pet) Ular Sanca Permata 100 Papua 7 Simalia (Morelia) tracyae 50 45 Total Halmahera python 50 Maluku Utara Hidup (Pet) Ular Sanca Halmahera (Halmahera) 8 Simalia (Morelia) nauta 50 45 Maluku Total Nauta Tanimbar Python 50 (yamdena dan Hidup (Pet) Ular Sanca Tanimbar Tanimbar) 9 Simalia (Morelia) clastolepis 50 45 maluku Total Southern Moluccas Python 50 Hidup (Pet) 1 Jatah Lokasi No. Nama Jenis Keterangan Tangkap Export Tangkap/Ambil Ular Sanca Seram (ambon & Seram) 10 Liasis mackloti mackoti 100 90 Total Macklot's Python / Ular -

Final Report (Phase : March 2008-February 2009)
Final Report (Phase : March 2008-February 2009) A Global Partnership for Community-Based Environmental Conservation in Salak Mountain : From Community Engagement to the Deepening Benefits of Forest Conservation Conducted By Funded By Peka Indonesia - Wildlife Trust Chevron Corporate Printed on recycle paper Content Summary............................................................................................................................................1 Chapter I : Introduction.....................................................................................................................2 1.1 Rationale ................................................................................................................................2 1.2 Historical Background............................................................................................................3 1.3 Goal and Objectives ...............................................................................................................4 Chapter II : Project Design and Strategy...........................................................................................5 2.1 Project Preparation................................................................................................................5 2.2 Program Design and Schedule...............................................................................................5 Chapter III : Inititave for Collaboration Management.....................................................................10 Chapter IV : Biodiversity -

Bugs R All Oct 2004.Pdf
Newsletter of the Invertebrate Conservation & Information Network of South Asia & the Invertebrate Special Interest Group, CBSG, South Asia Vol. 7 No. 2, October 2004 First National Stakeholders Consultation Workshop “CONSERVATION AND MANAGEMENT OF POLLINATORS FOR SUSTAINABLE AGRICULTURE, THROUGH AN ECOSYSTEM APPROACH" G.B. Pant Intitute of Himalayan Environment and Development, Almora, Uttaranchal, India October 8-9, 2004. The Ministry of Environment and Forest, Government of India, identified the G.B. Pant Institute of Himalayan Environ- ment and Development (GBPIHED), as the national executing agency for the PDF-B project (Project Development Facility-B), project on “Consultation workshop for conservation and Management of pollinators for sustainable Agri- culture, through ecosystem approach” that is being implemented by UNEP, approved by GEF and executed by FAO. The institute, under the leadership of Dr. Uppeandra Dhar, Director, GBPIHED organized the first National Stakehold- ers Consultation meeting during October 8-9, 2004 in Almora. The objective of the PDF-B project is to further the understanding of the issues related to the decline of pollinators that will lay the basis for activities in the Full-size project, and to design the full size project. The preparatory requirements to achieve this will focus on information collection, and analysis, establishment of local/national/re- gional/global priorities, and development of implementation strategies. Participating countries (Brazil, Ghana, India, Kenya, Nepal, Pakistan, and South Africa) and other stakeholders in the Full-size Project will be identified, collabo- rative effects will be established, and the donor co-financing will be mobilized during the PDF-B stage. This project aims to provide the added global benefits of information exchange, the dissemination of good practice, capacity building and exchanging knowledge at the farm, country, regional and global levels. -

Studies on the Taxonomy of Strongyloides (Nematoda; Strongyloididae)
ResearchOnline@JCU This file is part of the following work: Speare, Richard (1986) Studies on the taxonomy of Strongyloides (Nematoda; Strongyloididae). PhD thesis, James Cook University. Access to this file is available from: https://doi.org/10.25903/5e1bf2068703b Copyright © 1986 Richard Speare. If you believe that this work constitutes a copyright infringement, please email [email protected] STUDIES ON THE TAXONOMY OF STRONGYLOIDES (NEMATODA; STRONGYLOIDIDAE) Thesis submitted by Richard Speare, B.V.Sc., M.B., B.S. University of Queensland, St Lucia, Queensland for the Degree of Doctor of Philosophy in the Graduate School of Tropical Veterinary Science James Cook University of North Queensland March 1986 DECLARATION I declare that this thesis is ray own work and has not been submitted in any form for another degree or diploma at any university or other institute of tertiary education. Information derived from the published or unpublished work of others has been acknowledged in the text and a list of references is given. Richard Speare March, 1986 I, the undersigned, the author of this thesis, understand that the James Cook University of North Queensland will make it available for use within the University Library, and, by microfilm or other photographic means, allow access to users in other libraries. All users consulting this thesis will have to sign the following statement; "In consulting this thesis I agree to make proper written acknowledgement for any assistance which I have obtained from it". Beyond this, I do not wish to place any restriction on access to this thesis. In particular I wish to emphasise that I have no objection to this thesis being copied in whole or in part without my permission, and I deny any institution or organisation the right to impose this restriction on those wishing to do so.