Modafinil: a Review of Neurochemical Actions and Effects on Cognition
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Modafinil Tablets
PRODUCT MONOGRAPH INCLUDING PATIENT MEDICATION INFORMATION PrAURO-MODAFINIL Modafinil Tablets 100 mg House Standard Central Nervous System Stimulant Auro Pharma Inc. 3700 Steeles Avenue West, Suite # 402 Date of Revision: Woodbridge, ON, L4L 8K8, August 8, 2019. CANADA Submission Control Number: 230314 Page 1 of 41 Table of Contents PART I: HEALTH PROFESSIONAL INFORMATION ......................................................... 3 SUMMARY PRODUCT INFORMATION .................................................................... 3 INDICATIONS AND CLINICAL USE .......................................................................... 3 CONTRAINDICATIONS ............................................................................................... 4 WARNINGS AND PRECAUTIONS .............................................................................. 4 ADVERSE REACTIONS .............................................................................................. 12 DRUG INTERACTIONS .............................................................................................. 16 DOSAGE AND ADMINISTRATION .......................................................................... 19 OVERDOSAGE ............................................................................................................ 21 ACTION AND CLINICAL PHARMACOLOGY ........................................................ 21 STORAGE AND STABILITY ...................................................................................... 23 DOSAGE FORMS, COMPOSITION AND PACKAGING -
![[123I]FP-CIT SPECT in Atypical Degenerative Parkinsonism](https://docslib.b-cdn.net/cover/2351/123i-fp-cit-spect-in-atypical-degenerative-parkinsonism-242351.webp)
[123I]FP-CIT SPECT in Atypical Degenerative Parkinsonism
CONTRAST AGENT EVALUATION [123I]FP-CIT SPECT in atypical degenerative parkinsonism One of the most widely used techniques to support the clinical diagnosis of Parkinson’s disease is the SPECT scan with [123I]FP-CIT. This tracer binds reversibly and visualizes the striatal presynaptic dopamine transporters. Several uncertainties remain on the value of [123I]FP-CIT and SPECT in atypical degenerative parkinsonian syndromes. In this concise review, we discuss the contribution of SPECT and [123I]FP-CIT in supporting the clinical diagnosis of Parkinson’s disease and their role in the differential diagnosis of Parkinson’s disease and atypical degenerative parkinsonism. The chemistry, pharmacodynamics and pharmacokinetics of [123I]FP-CIT are also discussed. 1,2,3 KEywordS: atypical degenerative parkinsonism n FP-CIT n ioflupane n SPECT Ioannis U Isaias* , Giorgio Marotta4, Gianni Pezzoli2, Parkinson’s disease (PD) is the second most dystonic tremor [15] and psychogenic parkin- Osama Sabri5 [1] [16,17] 5,6 common neurodegenerative disorder , yet sonism . In this concise review, we will & Swen Hesse 123 early accurate diagnosis remains challenging. discuss the role of SPECT and [ I]FP-CIT in 1Università degli Studi di Milano, The estimated prevalence of PD is 0.5–1% in supporting the clinical diagnosis of PD and its Dipartimento di Fisiologia Umana, those aged 65–69 years and 1–3% in those aged differential diagnosis with ADP. Milano, Italy 2Centro per la Malattia di Parkinson e i ≥80 years [1]. Although the clinical diagnosis of Disturbi del Movimento, -

Modafinil and Modafinil Analogues: Free Radical Mechanism of the Eugeroic and Cognitive Enhancment Effect Clifford Fong
Modafinil and modafinil analogues: free radical mechanism of the eugeroic and cognitive enhancment effect Clifford Fong To cite this version: Clifford Fong. Modafinil and modafinil analogues: free radical mechanism of the eugeroic and cognitive enhancment effect. [Research Report] Eigenenergy. 2018. hal-01933737 HAL Id: hal-01933737 https://hal.archives-ouvertes.fr/hal-01933737 Submitted on 24 Nov 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Modafinil and modafinil analogues: free radical mechanism of the eugeroic and cognitive enhancment effect Clifford W. Fong Eigenenergy, Adelaide, South Australia. Keywords: Modafinil, modafinil-like analogues, eugeroic effect, cognitive enhancement, free radicals, quantum mechanics Abbreviations Dopamine DA, dopamine transporter DAT, Dissociative electron transfer or attachment DET, Linear free energy relationship LFER, free energy of water desolvation ΔG desolv,CDS , lipophilicity free energy ΔG lipo,CDS, cavity dispersion solvent structure of the first solvation shell CDS, highest occupied molecular orbital HOMO, lowest unoccupied molecular orbital LUMO, multiple correlation coefficient R 2, the F test of significance, standards errors for the estimate (SEE) and standard errors of the variables SE(ΔG desolCDS ), SE(ΔG lipoCDS ), SE(Dipole Moment), SE (Molecular Volume), transition state TS, reactive oxygen species ROS. -

These Highlights Do Not Include All the Information Needed to Use PROVIGIL Safely and Effectively
PROVIGIL- modafinil tablet Bryant Ranch Prepack ---------- HIGHLIGHTS OF PRESCRIBING INFORMATION These highlights do not include all the information needed to use PROVIGIL safely and effectively. See full prescribing information for PROVIGIL. PROVIGIL® (modafinil) tablets, for oral use, C-IV Initial U.S. Approval: 1998 INDICATIONS AND USAGE PROVIGIL is indicated to improve wakefulness in adult patients with excessive sleepiness associated with narcolepsy, obstructive sleep apnea (OSA), or shift work disorder (SWD). (1) Limitations of Use In OSA, PROVIGIL is indicated to treat excessive sleepiness and not as treatment for the underlying obstruction. DOSAGE AND ADMINISTRATION The recommended dosage of PROVIGIL for each indication is as follows: • Narcolepsy or OSA: 200 mg once a day in the morning. (2.1) • SWD: 200 mg once a day, taken approximately one hour prior to start of the work shift. (2.2) • Severe Hepatic Impairment: reduce dose to half the recommended dose. (2.3, 12.3) • Geriatric Patients: consider lower dose. (2.4, 12.3) DOSAGE FORMS AND STRENGTHS Tablets: 100 mg and 200 mg. (3) CONTRAINDICATIONS PROVIGIL is contraindicated in patients with known hypersensitivity to modafinil or armodafinil. (4) WARNINGS AND PRECAUTIONS • Serious Rash, including Stevens-Johnson Syndrome: Discontinue PROVIGIL at the first sign of rash, unless the rash is clearly not drug-related. (5.1) • Angioedema and Anaphylaxis Reactions: If suspected, discontinue PROVIGIL. (5.2) • Multi-organ Hypersensitivity Reactions: If suspected, discontinue PROVIGIL. (5.3) • Persistent Sleepiness: Assess patients frequently for degree of sleepiness and, if appropriate, advise patients to avoid driving or engaging in any other potentially dangerous activity. (5.4) • Psychiatric Symptoms: Use caution in patients with a history of psychosis, depression, or mania. -

A User's Guide to Methamphetamine
A USER’S GUIDE TO METHAMPHETAMINE A self-help guide to reduce harm for people who use methamphetamine 1st Edition, March 2017 Acknowledgements This booklet was adapted from an original publication created by The National Drug and Alcohol Research Centre, University of New South Wales, Sydney, Australia. This information does not constitute medical advice. Please seek the immediate help of a qualified medical practitioner about any personal health concerns. This booklet is being distributed for information purposes only. In the current state of crisis related to crystal methamphetamine, this booklet is intended as a guide to reduce harm for people who use methamphetamine. It lists the most common features of methamphetamine use, ways to reduce harm associated with the use of meth, and strategies for cutting down and quitting. The best way to avoid problems with drugs is to not use them. We are grateful for the contributions of the Integrated Drug Strategies in Waterloo Region and Guelph Wellington, in particular the leadership of Adrienne Crowder and Lindsay Sprague. Don Roth, Kerry Manthenga, Shirley Hilton, and our community review team provided great support and helpful edits. Marcey Gray provided exemplary skill on the design, images and editing, with final expert assistance from Arkay Design and Print. We are thankful for the financial support to print copies from the Waterloo-Wellington Human Services and Justice Coordinating Committee. For more information please contact: Wellington Guelph Drug Strategy www.wgdrugstrategy.ca Waterloo Region Integrated Drugs Strategy www.waterlooregiondrugstrategy.ca Circulated with the support of: WaterlooRegion Integrated Drugs Strategy WaterlooRegion Integrated Drugs Strategy The best way to avoid problems with drugs is to not use them. -

Contribution of Mass Spectrometry for the Detection of Xenobiotics
Contribution of mass spectrometry for the detection of xenobiotics implicated in cases of drug-facilitated crimes and the quantitation of urinary metabolites of polycyclic aromatic hydrocarbons Mohammed K.S. Shbair To cite this version: Mohammed K.S. Shbair. Contribution of mass spectrometry for the detection of xenobiotics implicated in cases of drug-facilitated crimes and the quantitation of urinary metabolites of polycyclic aromatic hydrocarbons. Human health and pathology. Université du Droit et de la Santé - Lille II, 2011. English. NNT : 2011LIL2S011. tel-00647316 HAL Id: tel-00647316 https://tel.archives-ouvertes.fr/tel-00647316 Submitted on 1 Dec 2011 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. University Lille 2 of Health and Law Doctorate School of Health and Biology Thesis for the Degree of Doctorate of Lille 2 University Discipline: TOXICOLOGY By Mohammed K.S. SHBAIR Contribution of mass spectrometry for the detection of xenobiotics implicated in cases of drug-facilitated crimes and the quantitation of urinary metabolites of polycyclic aromatic hydrocarbons Thesis defended 23rd June 2011 Members of the jury: Reviewers: Prof. Dr. Jean-Pierre GOULLÉ Prof. Dr. Christian STAUB Examiners : Prof. Dr. -

Texas Controlled Substances Act
HEALTH AND SAFETY CODE TITLE 6. FOOD, DRUGS, ALCOHOL, AND HAZARDOUS SUBSTANCES SUBTITLE C. SUBSTANCE ABUSE REGULATION AND CRIMES CHAPTER 481. TEXAS CONTROLLED SUBSTANCES ACT SUBCHAPTER A. GENERAL PROVISIONS Sec.A481.001.AASHORT TITLE. This chapter may be cited as the Texas Controlled Substances Act. Acts 1989, 71st Leg., ch. 678, Sec. 1, eff. Sept. 1, 1989. Sec.A481.002.AADEFINITIONS. In this chapter: (1)AA"Administer" means to directly apply a controlled substance by injection, inhalation, ingestion, or other means to the body of a patient or research subject by: (A)AAa practitioner or an agent of the practitioner in the presence of the practitioner; or (B)AAthe patient or research subject at the direction and in the presence of a practitioner. (2)AA"Agent" means an authorized person who acts on behalf of or at the direction of a manufacturer, distributor, or dispenser. The term does not include a common or contract carrier, public warehouseman, or employee of a carrier or warehouseman acting in the usual and lawful course of employment. (3)AA"Commissioner" means the commissioner of state health services or the commissioner 's designee. (4)AA"Controlled premises" means: (A)AAa place where original or other records or documents required under this chapter are kept or are required to be kept; or (B)AAa place, including a factory, warehouse, other establishment, or conveyance, where a person registered under this chapter may lawfully hold, manufacture, distribute, dispense, administer, possess, or otherwise dispose of a controlled substance or other item governed by the federal Controlled Substances Act (21 U.S.C. -

Pharmacy and Poisons Act 1979
Q UO N T FA R U T A F E BERMUDA PHARMACY AND POISONS ACT 1979 1979 : 26 TABLE OF CONTENTS PART I PRELIMINARY 1 Short title 2 Interpretation PART II THE PHARMACY COUNCIL 3 The Pharmacy Council 4 Membership of the Council 4A Functions of the Council 4B Protection from personal liability 4C Annual Report 5 Proceedings of the Council, etc PART III REGISTRATION OF PHARMACISTS 6 Offence to practise pharmacy if not registered 7 Registration as a pharmacist 7A Re-registration as non-practising member 7AA Period of validity of registration 8 Code of Conduct 9 Pharmacy Profession Complaints Committee 10 Investigation of complaint by Committee 10A Inquiry into complaint by Council 10B Inquiry by Council of its own initiative 11 Surrender of registration 12 Restoration of name to register 1 PHARMACY AND POISONS ACT 1979 13 Proof of registration 14 Appeals 14A Fees 14B Amendment of Seventh Schedule 15 Regulations for this part PART IV REGISTRATION OF PHARMACIES 16 Register of pharmacies 17 Registration of premises as registered pharmacies 18 Unfit premises: new applications 19 Unfit premises: registered pharmacies 20 Appeals 21 When certificates of unfitness take effect 22 Regulations for this Part PART V CONTROL OF PRESCRIPTIONS AND IMPORTATION 23 Prescriptions to be in a certain form 23A Validity of a prescription 24 Supply by registered pharmacist of equivalent medicines 25 Restrictions on the importation of medicines 26 Declaration relating to imported medicines [repealed] PART VI CONTROL OF DRUGS 27 Certain substances to be sold on prescription -

Alcohol Mixed with Other Drugs
Alcohol Mixed with Other Drugs Stimulants Stimulants or “uppers”: Drugs that temporarily +increase alertness and energy Examples: Adderall, Ritalin, cocaine, methamphetamine FOCUS: Alcohol + Adderall Adderall: Used to treat ADHD and narcolepsy. Some students misuse Adderall in hopes it will help them study. “Misuse” is defined as taking a medication that was not prescribed to you, taking more Alcohol than what was prescribed to you or taking it for a dierent purpose than prescribed. Eects: Because alcohol is a depressant Use CUPS to remember the and Adderall is a stimulant, Adderall will symptoms of alcohol poisoning: mask alcohol’s eects. Mixing alcohol with • Cold, clammy, pale or bluish skin. a stimulant makes you less aware of alcohol's • Unconscious or unable to be roused. intoxicating eects, which can result in an • Puking repeatedly or uncontrollably. overdose or death. Additionally, mixing alcohol with Adderall (or any other stimulant) • Slow or irregular breathing. can cause an irregular heartbeat and cause cardiovascular complications. Stat: 4.3 percent of UC Davis undergraduates reported using a prescription stimulant in the last 12 months that was not prescribed to them. Sedative Depressants or “downers”: Sedating drugs +that reduce stimulation Examples: opiates, Xanax, Valium FOCUS: Alcohol + Opiates/Opioids Opiates: A group of drugs that are used for treating pain -examples: heroin, morphine, codeine, oxycontin, vicodin, fentanyl Alcohol Eects: When alcohol and opioids are Symptoms: taken at the same time, the sedative • Slow or irregular breathing eects of both drugs will magnify. This • Lowered pulse and can depress or even stop involuntary blood pressure functions, such as breathing, and will • Unconscious or unable increase the risk of overdose and death. -

3514-18 Dowvigil P-I.FH10
160 mm 01 ® of expression of CYP2C9 activity. Other CYP activities may not appear to be affected 100mg Tablets by modafinil. Dowvigil 200mg Tablets Potential Interactions with Drugs that Inhibit, Induce, or are Metabolized by (Modafinil) Cytochrome P450 Isoenzymes and Other Hepatic Enzymes There may be a low probability of substantive effects on the overall pharmacokinetic profile of modafinil due to CYP inhibition by concomitant medications. Due to the partial DESCRIPTION involvement of CYP3A enzymes in the metabolic elimination of modafinil, coadministration Dowvigil (modafinil) is a wakefulness promoting agent for oral administration. Modafinil of potent inducers of CYP3A4/5 (e.g., carbamazepine, phenobarbital, and rifampin) or is a racemic compound. The chemical name for modafinil is 2-[(diphenylmethyl) inhibitors of CYP3A4/5 (e.g., ketoconazole, erythromycin) may alter the plasma sulfinyl]acetamide. The molecular formula is C15H15NO2S and the molecular weight is concentrations of modafinil. 273.35. The Potential of Modafinil to Alter the Metabolism of Other Drugs by Enzyme Induction or Inhibition COMPOSITION Drugs Metabolized by CYP3A4/5 Each tablet contains: Modafinil is a weak inducer of CYP3A activity in a concentration-related manner. Modafinil (USP)……………………………… 100mg Therefore, the blood levels and effectiveness of drugs that are substrates for CYP3A enzymes (e.g., steroidal contraceptives, cyclosporine, midazolam, and triazolam) may Each tablet contains: be reduced after initiation of concomitant treatment with modafinil. Modafinil (USP)……...……………………… 200mg Ethinyl Estradiol: Administration of modafinil once daily at 200mg/day for 7 days followed by 400mg/day for 21 days may result in a mean 11% decrease in mean Cmax THERAPEUTIC INDICATIONS and 18% decrease in mean AUC0-24 of ethinyl estradiol.There may be no apparent Dowvigil is indicated to improve wakefulness in adult individuals with excessive change in the elimination rate of ethinyl estradiol. -
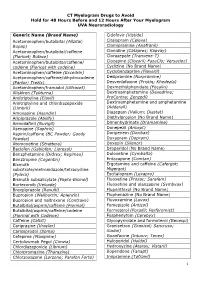
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem) -

Prescription Stimulants
Prescription Stimulants What are prescription stimulants? Prescription stimulants are medicines generally used to treat attention-deficit hyperactivity disorder (ADHD) and narcolepsy— uncontrollable episodes of deep sleep. They increase alertness, attention, and energy. What are common prescription stimulants? • dextroamphetamine (Dexedrine®) • dextroamphetamine/amphetamine combination product (Adderall®) • methylphenidate (Ritalin®, Concerta®). Photo by ©iStock.com/ognianm Popular slang terms for prescription stimulants include Speed, Uppers, and Vitamin R. How do people use and misuse prescription stimulants? Most prescription stimulants come in tablet, capsule, or liquid form, which a person takes by mouth. Misuse of a prescription stimulant means: Do Prescription Stimulants Make You • taking medicine in a way or dose Smarter? other than prescribed Some people take prescription stimulants to • taking someone else’s medicine try to improve mental performance. Teens • taking medicine only for the effect it and college students sometimes misuse causes—to get high them to try to get better grades, and older adults misuse them to try to improve their When misusing a prescription stimulant, memory. Taking prescription stimulants for people can swallow the medicine in its reasons other than treating ADHD or normal form. Alternatively, they can crush narcolepsy could lead to harmful health tablets or open the capsules, dissolve the powder in water, and inject the liquid into a effects, such as addiction, heart problems, vein. Some can also snort or smoke the or psychosis. powder. Prescription Stimulants • June 2018 • Page 1 How do prescription stimulants affect the brain and body? Prescription stimulants increase the activity of the brain chemicals dopamine and norepinephrine. Dopamine is involved in the reinforcement of rewarding behaviors.