Redalyc.Geographic Distribution and Description of Four Pelagic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Cirripedia : Thoracica) Over the Body of a Sea Snake, Laticauda Title Semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan
Distribution of Two Species of Conchoderma (Cirripedia : Thoracica) over the Body of a Sea Snake, Laticauda Title semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan Yamato, Shigeyuki; Yusa, Yoichi; Tanase, Hidetomo; Tanase, Author(s) Hidetomo PUBLICATIONS OF THE SETO MARINE BIOLOGICAL Citation LABORATORY (1996), 37(3-6): 337-343 Issue Date 1996-12-25 URL http://hdl.handle.net/2433/176259 Right Type Departmental Bulletin Paper Textversion publisher Kyoto University Pub!. Seto Mar. Bioi. Lab., 37(3/6): 337-343, 1996 337 Distribution of Two Species of Conchoderma (Cirripedia: Thoracica) over the Body of a Sea Snake, Laticauda semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan SHIGEYUKI YAMATO, YOICHI YUSA and HIDETOMO TANASE Seto Marine Biological Laboratory, Kyoto University, Shirahama, Wakayama 649-22, Japan Abstract Two species of Conchoderma were found on a sea snake, Laticauda semifas ciata (Reinwardt), collected on the west coast of the Kii Peninsula. A total of 223 individuals of C. virgatum and 6 of C. hunteri in 19 clumps were attached to the snake's body. The barnacles ranged in size from 1.4 mm (cypris larvae) to 18.2 mm in capitulum length in C. virgatum, and from 10.7 to 14.4 mm in C. hunteri. The size of the smallest gravid individuals in both species was between 10 and 11 mm. The distribution of C. virgatum on the snake was non-random both longitudinally and dorso-ventrally, with more barnacles in the posterior region and on the ventral side of the snake, respectively. The proportion of gravid individuals increased towards the tail. -
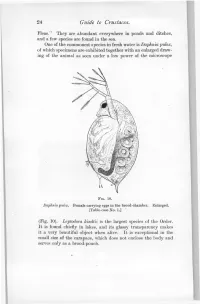
24 Guide to Crustacea
24 Guide to Crustacea. Fleas." They are abundant everywhere in ponds and ditches, and a few species are found in the sea. One of the commonest species in fresh water is Daphnia pulex, of which specimens are exhibited together with an enlarged draw- ing of the animal as seen under a low power of the microscope FIG. 10. Daphnia pulex. Female carrying eggs in the brood-chamber. Enlarged. [Table-case No. 1.] (Fig. 10). Leptodora kindtii is the largest species of the Order. It is found chiefly in lakes, and its glassy transparency makes it a very beautiful object when alive. It is exceptional in the small size of the carapace, which does not enclose the body and serves only as a brood-pouch. Ostracoda. 25 Sub-class II.—OSTRACODA. (Table-ease No. 1.) The number of somites, as indicated by the appendages, is smaller than in any other Crustacea, there being, at most, only two pairs of trunk-limbs behind those belonging to the head- region. The carapace forms a bivalved shell completely en- closing the body and limbs. There is a large, and often leg-like, palp on the mandible. The antennules and antennae are used for creeping or swimming. The Ostracoda (Fig. 10) are for the most part extremely minute animals, and only one or two of the larger species can be exhibited. They occur abundantly in fresh water and in FIG. 11. Shells of Ostracoda, seen from the side. A. Philomedes brenda (Myodocopa) ; B. Cypris fuscata (Podocopa); ('. Cythereis ornata (Podocopa): all much enlarged, n., Notch characteristic of the Myodocopa; e., the median eye ; a., mark of attachment of the muscle connecting the two valves of the shell. -

Balanus Glandula Class: Multicrustacea, Hexanauplia, Thecostraca, Cirripedia
Phylum: Arthropoda, Crustacea Balanus glandula Class: Multicrustacea, Hexanauplia, Thecostraca, Cirripedia Order: Thoracica, Sessilia, Balanomorpha Acorn barnacle Family: Balanoidea, Balanidae, Balaninae Description (the plate overlapping plate edges) and radii Size: Up to 3 cm in diameter, but usually (the plate edge marked off from the parietes less than 1.5 cm (Ricketts and Calvin 1971; by a definite change in direction of growth Kozloff 1993). lines) (Fig. 3b) (Newman 2007). The plates Color: Shell usually white, often irregular themselves include the carina, the carinola- and color varies with state of erosion. Cirri teral plates and the compound rostrum (Fig. are black and white (see Plate 11, Kozloff 3). 1993). Opercular Valves: Valves consist of General Morphology: Members of the Cirri- two pairs of movable plates inside the wall, pedia, or barnacles, can be recognized by which close the aperture: the tergum and the their feathery thoracic limbs (called cirri) that scutum (Figs. 3a, 4, 5). are used for feeding. There are six pairs of Scuta: The scuta have pits on cirri in B. glandula (Fig. 1). Sessile barna- either side of a short adductor ridge (Fig. 5), cles are surrounded by a shell that is com- fine growth ridges, and a prominent articular posed of a flat basis attached to the sub- ridge. stratum, a wall formed by several articulated Terga: The terga are the upper, plates (six in Balanus species, Fig. 3) and smaller plate pair and each tergum has a movable opercular valves including terga short spur at its base (Fig. 4), deep crests for and scuta (Newman 2007) (Figs. -

Dosima Fascicularis (Ellis & Solander, 1786) (Crustacea, Cirripedia) from the Galician Istributio D
Check List 10(3): 669–671, 2014 © 2014 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution N First record of the buoy barnacle Dosima fascicularis (Ellis & Solander, 1786) (Crustacea, Cirripedia) from the Galician ISTRIBUTIO D * RAPHIC G beaches (NW Spain) after the Prestige oil spill EO G Juan Junoy and Jorge Junoy N O EU-US Marine Biodiversity [email protected] Group, Franklin Institute and Departamento de Ciencias de la Vida, AP 20. Campus Universitario, Universidad de Alcalá, 28805 Alcalá de Henares, Spain. OTES * Corresponding author. E-mail: N Abstract: Dosima fascicularis The presence of the buoy barnacle is first documented for Spanish waters. More than the half of the specimens collected was attached to tar pellets from the Prestige oil spill. DOI: 10.15560/10.3.669 Dosima fascicularis (Ellis & Solander, 1786) is a presumablycollected along from the the 1470 Prestige m of oil the spill; beach other longitude. attachment The stalked barnacle which capitulum bears five large plates. surfacesfloats of 53.8%were Velellaof the specimens were Sargassumattached to bladderstar balls, It is the unique barnacle that constructs its own float, a attachmentfoamy mass surfaces described (Darwin as similar 1851; to Minchin polystyrene, 1996; usingRyan tests (15.3%), floating items as feathers, tar balls and plastic particles as (7.6%) and feathers (2.5%). 20.8% of buoy barnacles lacked an obvious attachment object (Figure 2). theand BritishBranch Islands2012). The and species Baltic Seais widespread (Darwin 1851) in temperate but its sizeThe recorded average for thecapitulum species lengthwas 37 ofmm the in Europeancollected presenceand subtropical in European ocean waters,coast is reaching unusual in(O’ the Riodan NE Atlantic 1967; specimens was 17.7 mm (range 13–22 mm). -

A Checklist of Turtle and Whale Barnacles
Journal of the Marine Biological Association of the United Kingdom, 2013, 93(1), 143–182. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S0025315412000847 A checklist of turtle and whale barnacles (Cirripedia: Thoracica: Coronuloidea) ryota hayashi1,2 1International Coastal Research Center, Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5, Kashiwanoha, Kashiwa-shi, Chiba 277-8564 Japan, 2Marine Biology and Ecology Research Program, Extremobiosphere Research Center, Japan Agency for Marine–Earth Science and Technology A checklist of published records of coronuloid barnacles (Cirripedia: Thoracica: Coronuloidea) attached to marine vertebrates is presented, with 44 species (including 15 fossil species) belonging to 14 genera (including 3 fossil genera) and 3 families recorded. Also included is information on their geographical distribution and the hosts with which they occur. Keywords: checklist, turtle barnacles, whale barnacles, Chelonibiidae, Emersoniidae, Coronulidae, Platylepadidae, host and distribution Submitted 10 May 2012; accepted 16 May 2012; first published online 10 August 2012 INTRODUCTION Superorder THORACICA Darwin, 1854 Order SESSILIA Lamarck, 1818 In this paper, a checklist of barnacles of the superfamily Suborder BALANOMORPHA Pilsbry, 1916 Coronuloidea occurring on marine animals is presented. Superfamily CORONULOIDEA Newman & Ross, 1976 The systematic arrangement used herein follows Newman Family CHELONIBIIDAE Pilsbry, 1916 (1996) rather than Ross & Frick (2011) for reasons taken up in Hayashi (2012) in some detail. The present author Genus Chelonibia Leach, 1817 deems the subfamilies of the Cheonibiidae (Chelonibiinae, Chelonibia caretta (Spengler, 1790) Emersoniinae and Protochelonibiinae) proposed by Harzhauser et al. (2011), as well as those included of Ross & Lepas caretta Spengler, 1790: 185, plate 6, figure 5. -

Remarkable Convergent Evolution in Specialized Parasitic Thecostraca (Crustacea)
Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea) Pérez-Losada, Marcos; Høeg, Jens Thorvald; Crandall, Keith A Published in: BMC Biology DOI: 10.1186/1741-7007-7-15 Publication date: 2009 Document version Publisher's PDF, also known as Version of record Citation for published version (APA): Pérez-Losada, M., Høeg, J. T., & Crandall, K. A. (2009). Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea). BMC Biology, 7(15), 1-12. https://doi.org/10.1186/1741-7007-7-15 Download date: 25. Sep. 2021 BMC Biology BioMed Central Research article Open Access Remarkable convergent evolution in specialized parasitic Thecostraca (Crustacea) Marcos Pérez-Losada*1, JensTHøeg2 and Keith A Crandall3 Address: 1CIBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Universidade do Porto, Campus Agrário de Vairão, Portugal, 2Comparative Zoology, Department of Biology, University of Copenhagen, Copenhagen, Denmark and 3Department of Biology and Monte L Bean Life Science Museum, Brigham Young University, Provo, Utah, USA Email: Marcos Pérez-Losada* - [email protected]; Jens T Høeg - [email protected]; Keith A Crandall - [email protected] * Corresponding author Published: 17 April 2009 Received: 10 December 2008 Accepted: 17 April 2009 BMC Biology 2009, 7:15 doi:10.1186/1741-7007-7-15 This article is available from: http://www.biomedcentral.com/1741-7007/7/15 © 2009 Pérez-Losada et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. -

A New Species of Lepas (Crustacea: Cirripedia: Pedunculata) from the Miocene Mizunami Group, Japan
Bulletin of the Mizunami Fossil Museum, no. 31 (2004), p. 91-93, 1 fig. c 2004, Mizunami Fossil Museum A new species of Lepas (Crustacea: Cirripedia: Pedunculata) from the Miocene Mizunami Group, Japan Hiroaki Karasawa*, Toshio Tanaka* *, and Yoshitsugu Okumura*** * Mizunami Fossil Museum, Yamanouchi, Akeyo, Mizunami, Gifu 509-6132, Japan <[email protected]> ** Aichi Gakuin Junior College, Chikusa, Nagoya 464-8650, Japan < [email protected]> *** Mizunami Fossil Museum, Yamanouchi, Akeyo, Mizunami, Gifu 509-6132, Japan <[email protected]> Abstract Lepas kuwayamai, a new species of a lepadid thracican is described from the lower Miocene Mizunami Group in Gifu Prefecture, Honshu, Japan. This represents the third record for the family from Miocene deposits of Japan. Key words: Crustacea, Cirripedia, Thoracica, Pedunculata, Lepadidae, Lepas, Miocene, Japan Material examined: MFM9043, holotype; 8 paratypes, Introduction MFM9044-9051; Coll. M. Kuwayama in, 2004. All specimens are housed in the Mizunami Fossil Museum. The lepadomorph family Lepadidae Darwin, 1851, is a Diagnosis: Lepas with moderate sized capitulum. Shell small group including six genera (Newman, 1996). Among thick. Scutum triangular, slightly higher than wide, with these, the genus Lepas Linnaeus, 1758, is only known clear growth lines; umbonal tooth absent; very weak from Japan in the fossil record. O’hara et al. (1976) reported radial striae sometimes present; apicoumbonal ridge Lepas sp. from the middle Pleistocene Shimosa Group and weak. Tergum flattened without radial striae; occludent unnamed Miocene species was reported from the lower margin convex, rounded. Carina broad. Miocene Morozaki Group (Mizuno and Takeda, 1993) and Etymology: In honor to our friend, Mr. -

An Invitation to Monitor Georgia's Coastal Wetlands
An Invitation to Monitor Georgia’s Coastal Wetlands www.shellfish.uga.edu By Mary Sweeney-Reeves, Dr. Alan Power, & Ellie Covington First Printing 2003, Second Printing 2006, Copyright University of Georgia “This book was prepared by Mary Sweeney-Reeves, Dr. Alan Power, and Ellie Covington under an award from the Office of Ocean and Coastal Resource Management, National Oceanic and Atmospheric Administration. The statements, findings, conclusions, and recommendations are those of the authors and do not necessarily reflect the views of OCRM and NOAA.” 2 Acknowledgements Funding for the development of the Coastal Georgia Adopt-A-Wetland Program was provided by a NOAA Coastal Incentive Grant, awarded under the Georgia Department of Natural Resources Coastal Zone Management Program (UGA Grant # 27 31 RE 337130). The Coastal Georgia Adopt-A-Wetland Program owes much of its success to the support, experience, and contributions of the following individuals: Dr. Randal Walker, Marie Scoggins, Dodie Thompson, Edith Schmidt, John Crawford, Dr. Mare Timmons, Marcy Mitchell, Pete Schlein, Sue Finkle, Jenny Makosky, Natasha Wampler, Molly Russell, Rebecca Green, and Jeanette Henderson (University of Georgia Marine Extension Service); Courtney Power (Chatham County Savannah Metropolitan Planning Commission); Dr. Joe Richardson (Savannah State University); Dr. Chandra Franklin (Savannah State University); Dr. Dionne Hoskins (NOAA); Dr. Charles Belin (Armstrong Atlantic University); Dr. Merryl Alber (University of Georgia); (Dr. Mac Rawson (Georgia Sea Grant College Program); Harold Harbert, Kim Morris-Zarneke, and Michele Droszcz (Georgia Adopt-A-Stream); Dorset Hurley and Aimee Gaddis (Sapelo Island National Estuarine Research Reserve); Dr. Charra Sweeney-Reeves (All About Pets); Captain Judy Helmey (Miss Judy Charters); Jan Mackinnon and Jill Huntington (Georgia Department of Natural Resources). -

An Annotated Checklist of the Marine Macroinvertebrates of Alaska David T
NOAA Professional Paper NMFS 19 An annotated checklist of the marine macroinvertebrates of Alaska David T. Drumm • Katherine P. Maslenikov Robert Van Syoc • James W. Orr • Robert R. Lauth Duane E. Stevenson • Theodore W. Pietsch November 2016 U.S. Department of Commerce NOAA Professional Penny Pritzker Secretary of Commerce National Oceanic Papers NMFS and Atmospheric Administration Kathryn D. Sullivan Scientific Editor* Administrator Richard Langton National Marine National Marine Fisheries Service Fisheries Service Northeast Fisheries Science Center Maine Field Station Eileen Sobeck 17 Godfrey Drive, Suite 1 Assistant Administrator Orono, Maine 04473 for Fisheries Associate Editor Kathryn Dennis National Marine Fisheries Service Office of Science and Technology Economics and Social Analysis Division 1845 Wasp Blvd., Bldg. 178 Honolulu, Hawaii 96818 Managing Editor Shelley Arenas National Marine Fisheries Service Scientific Publications Office 7600 Sand Point Way NE Seattle, Washington 98115 Editorial Committee Ann C. Matarese National Marine Fisheries Service James W. Orr National Marine Fisheries Service The NOAA Professional Paper NMFS (ISSN 1931-4590) series is pub- lished by the Scientific Publications Of- *Bruce Mundy (PIFSC) was Scientific Editor during the fice, National Marine Fisheries Service, scientific editing and preparation of this report. NOAA, 7600 Sand Point Way NE, Seattle, WA 98115. The Secretary of Commerce has The NOAA Professional Paper NMFS series carries peer-reviewed, lengthy original determined that the publication of research reports, taxonomic keys, species synopses, flora and fauna studies, and data- this series is necessary in the transac- intensive reports on investigations in fishery science, engineering, and economics. tion of the public business required by law of this Department. -

Barnacle Paper.PUB
Proc. Isle Wight nat. Hist. archaeol. Soc . 24 : 42-56. BARNACLES (CRUSTACEA: CIRRIPEDIA) OF THE SOLENT & ISLE OF WIGHT Dr Roger J.H. Herbert & Erik Muxagata To coincide with the bicentenary of the birth of the naturalist Charles Darwin (1809-1889) a list of barnacles (Crustacea:Cirripedia) recorded from around the Solent and Isle of Wight coast is pre- sented, including notes on their distribution. Following the Beagle expedition, and prior to the publication of his seminal work Origin of Species in 1859, Darwin spent eight years studying bar- nacles. During this time he tested his developing ideas of natural selection and evolution through precise observation and systematic recording of anatomical variation. To this day, his monographs of living and fossil cirripedia (Darwin 1851a, 1851b, 1854a, 1854b) are still valuable reference works. Darwin visited the Isle of Wight on three occasions (P. Bingham, pers.com) however it is unlikely he carried out any field work on the shore. He does however describe fossil cirripedia from Eocene strata on the Isle of Wight (Darwin 1851b, 1854b) and presented specimens, that were supplied to him by other collectors, to the Natural History Museum (Appendix). Barnacles can be the most numerous of macrobenthic species on hard substrata. The acorn and stalked (pedunculate) barnacles have a familiar sessile adult stage that is preceded by a planktonic larval phase comprising of six naupliar stages, prior to the metamorphosis of a non-feeding cypris that eventually settles on suitable substrate (for reviews on barnacle biology see Rainbow 1984; Anderson, 1994). Additionally, the Rhizocephalans, an ectoparasitic group, are mainly recognis- able as barnacles by the external characteristics of their planktonic nauplii. -

CIRRIPEDIA Sheet 83 Thoracica, Family: Lepadidae (By Hjalmar Broch) 1959
CONSEIL INTERNATIONAL POUR L'EXPLORATION DE LA MER Zooplankton CIRRIPEDIA Sheet 83 Thoracica, Family: Lepadidae (By Hjalmar Broch) 1959 https://doi.org/10.17895/ices.pub.5008 Comprises the region north of 40" N Lat. -2- -I -I I 4 P 111 2d 5c Ill - IV V 5a 5b 7 5d 1, Lepas anatifera. 2, L. hillil. :;, L. unteriferu. 4, L. pectinata. 5, L. fascicularis. 6, Conchodermu auritum. 7, C. virgatuna. a, side view; b, dorsal view; c, ventral view of the basi-scutal region; d, side view of the body showing filamentary appendages (I-V). (Fig. 3 redrawn after P i 1 s b r y, the other figures from B r o c h) Capitulum at most with 5 plates. Peduncle naked. Filamentary appendages at the hase of the cirri. Margin of the maxilla with step-formed edge . .. .. .. .. .. .. .. .. .. .. .. .. .. .. LEPADIDAE - 3 - Family LEPADIDAE Genus LEPAS Mn. Capitulum with 5 well developed plates separated by narrow interspaces. Apical part of carina between the terga. Subgenus Anatifa (Bruguiere): Carina basally ending in a fork at the sides of umbo embedded in the membrane just below the basal margin of the scuta. Subgenus Dosima (Gray): Carina with a great, almost rectangularly inflected disc below umbo, not embedded in the membrane below capitulum. Capitulum plates Species Peduncle Filamentary Capitulum appendages length Remarks Carina Scutum Tergum 1. Lepas Dark purplish Generally Smooth, seldom A little convex, 2, seldom 1, upto (Anati f a) brown, especially smooth, seldom with feeble obliquely on each side about anatifera dark just below barbed stripes radiating quadrangular 5 cm (Lin.) capitulum from the basi- (trapezoid) ventral umbo. -

Title FEEDING PREFERENCES and RATES of the SNAIL, IANTHINA
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Kyoto University Research Information Repository FEEDING PREFERENCES AND RATES OF THE SNAIL, IANTHINA PROLONGATA, THE BARNACLE, LEPAS Title ANSERIFERA, THE NUDIBRANCHS, GLAUCUS ATLANTICUS AND FIONA PINNATA, AND THE FOOD WEB IN THE MARINE NEUSTON Author(s) Bieri, Robert PUBLICATIONS OF THE SETO MARINE BIOLOGICAL Citation LABORATORY (1966), 14(2): 161-170 Issue Date 1966-06-30 URL http://hdl.handle.net/2433/175429 Right Type Departmental Bulletin Paper Textversion publisher Kyoto University FEEDING PREFERENCES AND RATES OF THE SNAIL, IANTHINA PROLONGATA, THE BARNACLE, LEPAS ANSERIFERA, THE NUDIBRANCHS, GLAUCUS ATLANTICUS AND FIONA PINNATA, AND THE FOOD WEB IN THE MARINE NEUSTON'l RoBERT BIERI Antioch College, Yellow Springs, Ohio With Plates III-IV and 1 Text-figure On the afternoon of November 5, 1965 during strong north-northwest winds, Porpita, Velella, Physalia, G!aucus, Lepas, Fiona, and three species of Ianthina were blown onto the north beach of the Seto Marine Biological Laboratory. In the Laboratory with an ample supply of running sea water, many of these animals lived for several days and I was able to observe their feeding preferences and rates. On the basis of these obervations and the reports of other workers it is possible to construct a preliminary diagram of the neuston food web. Ianthina prolongata Of the three species of Ianthina washed ashore, I. janthina forma balteata, I. umbilicata, and I. prolongata, I. prolongata was most abundant. I was able to collect more than 160 of them in less than an hour.