Title FEEDING PREFERENCES and RATES of the SNAIL, IANTHINA
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Appendix to Taxonomic Revision of Leopold and Rudolf Blaschkas' Glass Models of Invertebrates 1888 Catalogue, with Correction
http://www.natsca.org Journal of Natural Science Collections Title: Appendix to Taxonomic revision of Leopold and Rudolf Blaschkas’ Glass Models of Invertebrates 1888 Catalogue, with correction of authorities Author(s): Callaghan, E., Egger, B., Doyle, H., & E. G. Reynaud Source: Callaghan, E., Egger, B., Doyle, H., & E. G. Reynaud. (2020). Appendix to Taxonomic revision of Leopold and Rudolf Blaschkas’ Glass Models of Invertebrates 1888 Catalogue, with correction of authorities. Journal of Natural Science Collections, Volume 7, . URL: http://www.natsca.org/article/2587 NatSCA supports open access publication as part of its mission is to promote and support natural science collections. NatSCA uses the Creative Commons Attribution License (CCAL) http://creativecommons.org/licenses/by/2.5/ for all works we publish. Under CCAL authors retain ownership of the copyright for their article, but authors allow anyone to download, reuse, reprint, modify, distribute, and/or copy articles in NatSCA publications, so long as the original authors and source are cited. TABLE 3 – Callaghan et al. WARD AUTHORITY TAXONOMY ORIGINAL SPECIES NAME REVISED SPECIES NAME REVISED AUTHORITY N° (Ward Catalogue 1888) Coelenterata Anthozoa Alcyonaria 1 Alcyonium digitatum Linnaeus, 1758 2 Alcyonium palmatum Pallas, 1766 3 Alcyonium stellatum Milne-Edwards [?] Sarcophyton stellatum Kükenthal, 1910 4 Anthelia glauca Savigny Lamarck, 1816 5 Corallium rubrum Lamarck Linnaeus, 1758 6 Gorgonia verrucosa Pallas, 1766 [?] Eunicella verrucosa 7 Kophobelemon (Umbellularia) stelliferum -

15 Sea Turtle Epibiosis
15 Sea Turtle Epibiosis Michael G. Frick and joseph B. Pfaller CONTENTS 15. I Introduction .......................................................................................................................... 399 15.2 Common Forms .................................................................................................................... 401 15.2.1 Sessile Forms ............................................................................................................ 401 15.2.2 Sedentary Forms ....................................................................................................... 401 15.2.3 Motile Forms ............................................................................................................ 401 15.3 Communities and Community Dynamics ............................................................................ 402 15.3.1 Pelagic/Oceanic Communities .................................................................................. 402 15.3.2 Benthic/Neritic Communities ................................................................................... 402 15.3.3 Obligate Communities .............................................................................................. 403 15.3.4 Community Distribution ........................................................................................... 403 15.3.5 Community Succession ............................................................................................ 404 15.4 Ecological Interactions ........................................................................................................ -

The Evolution of Siphonophore Tentilla for Specialized Prey Capture in the Open Ocean
The evolution of siphonophore tentilla for specialized prey capture in the open ocean Alejandro Damian-Serranoa,1, Steven H. D. Haddockb,c, and Casey W. Dunna aDepartment of Ecology and Evolutionary Biology, Yale University, New Haven, CT 06520; bResearch Division, Monterey Bay Aquarium Research Institute, Moss Landing, CA 95039; and cEcology and Evolutionary Biology, University of California, Santa Cruz, CA 95064 Edited by Jeremy B. C. Jackson, American Museum of Natural History, New York, NY, and approved December 11, 2020 (received for review April 7, 2020) Predator specialization has often been considered an evolutionary makes them an ideal system to study the relationships between “dead end” due to the constraints associated with the evolution of functional traits and prey specialization. Like a head of coral, a si- morphological and functional optimizations throughout the organ- phonophore is a colony bearing many feeding polyps (Fig. 1). Each ism. However, in some predators, these changes are localized in sep- feeding polyp has a single tentacle, which branches into a series of arate structures dedicated to prey capture. One of the most extreme tentilla. Like other cnidarians, siphonophores capture prey with cases of this modularity can be observed in siphonophores, a clade of nematocysts, harpoon-like stinging capsules borne within special- pelagic colonial cnidarians that use tentilla (tentacle side branches ized cells known as cnidocytes. Unlike the prey-capture apparatus of armed with nematocysts) exclusively for prey capture. Here we study most other cnidarians, siphonophore tentacles carry their cnidocytes how siphonophore specialists and generalists evolve, and what mor- in extremely complex and organized batteries (3), which are located phological changes are associated with these transitions. -

A Radical Solution: the Phylogeny of the Nudibranch Family Fionidae
RESEARCH ARTICLE A Radical Solution: The Phylogeny of the Nudibranch Family Fionidae Kristen Cella1, Leila Carmona2*, Irina Ekimova3,4, Anton Chichvarkhin3,5, Dimitry Schepetov6, Terrence M. Gosliner1 1 Department of Invertebrate Zoology, California Academy of Sciences, San Francisco, California, United States of America, 2 Department of Marine Sciences, University of Gothenburg, Gothenburg, Sweden, 3 Far Eastern Federal University, Vladivostok, Russia, 4 Biological Faculty, Moscow State University, Moscow, Russia, 5 A.V. Zhirmunsky Instutute of Marine Biology, Russian Academy of Sciences, Vladivostok, Russia, 6 National Research University Higher School of Economics, Moscow, Russia a11111 * [email protected] Abstract Tergipedidae represents a diverse and successful group of aeolid nudibranchs, with approx- imately 200 species distributed throughout most marine ecosystems and spanning all bio- OPEN ACCESS geographical regions of the oceans. However, the systematics of this family remains poorly Citation: Cella K, Carmona L, Ekimova I, understood since no modern phylogenetic study has been undertaken to support any of the Chichvarkhin A, Schepetov D, Gosliner TM (2016) A Radical Solution: The Phylogeny of the proposed classifications. The present study is the first molecular phylogeny of Tergipedidae Nudibranch Family Fionidae. PLoS ONE 11(12): based on partial sequences of two mitochondrial (COI and 16S) genes and one nuclear e0167800. doi:10.1371/journal.pone.0167800 gene (H3). Maximum likelihood, maximum parsimony and Bayesian analysis were con- Editor: Geerat J. Vermeij, University of California, ducted in order to elucidate the systematics of this family. Our results do not recover the tra- UNITED STATES ditional Tergipedidae as monophyletic, since it belongs to a larger clade that includes the Received: July 7, 2016 families Eubranchidae, Fionidae and Calmidae. -
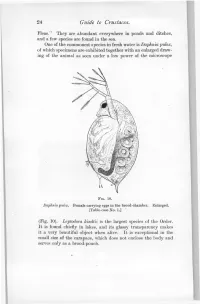
24 Guide to Crustacea
24 Guide to Crustacea. Fleas." They are abundant everywhere in ponds and ditches, and a few species are found in the sea. One of the commonest species in fresh water is Daphnia pulex, of which specimens are exhibited together with an enlarged draw- ing of the animal as seen under a low power of the microscope FIG. 10. Daphnia pulex. Female carrying eggs in the brood-chamber. Enlarged. [Table-case No. 1.] (Fig. 10). Leptodora kindtii is the largest species of the Order. It is found chiefly in lakes, and its glassy transparency makes it a very beautiful object when alive. It is exceptional in the small size of the carapace, which does not enclose the body and serves only as a brood-pouch. Ostracoda. 25 Sub-class II.—OSTRACODA. (Table-ease No. 1.) The number of somites, as indicated by the appendages, is smaller than in any other Crustacea, there being, at most, only two pairs of trunk-limbs behind those belonging to the head- region. The carapace forms a bivalved shell completely en- closing the body and limbs. There is a large, and often leg-like, palp on the mandible. The antennules and antennae are used for creeping or swimming. The Ostracoda (Fig. 10) are for the most part extremely minute animals, and only one or two of the larger species can be exhibited. They occur abundantly in fresh water and in FIG. 11. Shells of Ostracoda, seen from the side. A. Philomedes brenda (Myodocopa) ; B. Cypris fuscata (Podocopa); ('. Cythereis ornata (Podocopa): all much enlarged, n., Notch characteristic of the Myodocopa; e., the median eye ; a., mark of attachment of the muscle connecting the two valves of the shell. -

US Fish & Wildlife Service Seabird Conservation Plan—Pacific Region
U.S. Fish & Wildlife Service Seabird Conservation Plan Conservation Seabird Pacific Region U.S. Fish & Wildlife Service Seabird Conservation Plan—Pacific Region 120 0’0"E 140 0’0"E 160 0’0"E 180 0’0" 160 0’0"W 140 0’0"W 120 0’0"W 100 0’0"W RUSSIA CANADA 0’0"N 0’0"N 50 50 WA CHINA US Fish and Wildlife Service Pacific Region OR ID AN NV JAP CA H A 0’0"N I W 0’0"N 30 S A 30 N L I ort I Main Hawaiian Islands Commonwealth of the hwe A stern A (see inset below) Northern Mariana Islands Haw N aiian Isla D N nds S P a c i f i c Wake Atoll S ND ANA O c e a n LA RI IS Johnston Atoll MA Guam L I 0’0"N 0’0"N N 10 10 Kingman Reef E Palmyra Atoll I S 160 0’0"W 158 0’0"W 156 0’0"W L Howland Island Equator A M a i n H a w a i i a n I s l a n d s Baker Island Jarvis N P H O E N I X D IN D Island Kauai S 0’0"N ONE 0’0"N I S L A N D S 22 SI 22 A PAPUA NEW Niihau Oahu GUINEA Molokai Maui 0’0"S Lanai 0’0"S 10 AMERICAN P a c i f i c 10 Kahoolawe SAMOA O c e a n Hawaii 0’0"N 0’0"N 20 FIJI 20 AUSTRALIA 0 200 Miles 0 2,000 ES - OTS/FR Miles September 2003 160 0’0"W 158 0’0"W 156 0’0"W (800) 244-WILD http://www.fws.gov Information U.S. -

Parallel Changes in Genital Morphology Delineate Cryptic Diversification of Planktonic Nudibranchs
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Poet Commons (Whittier College) Whittier College Poet Commons Biology Faculty Publications & Research 2013 Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs Alvin Alejandrino [email protected] Celia K. C. Churchill Ángel Valdés Diarmaid Ó. Foighil Follow this and additional works at: https://poetcommons.whittier.edu/bio Part of the Biology Commons Recommended Citation Alejandrino, Alvin; Churchill, Celia K. C.; Valdés, Ángel; and Foighil, Diarmaid Ó., "Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs" (2013). Biology. 1. https://poetcommons.whittier.edu/bio/1 This Article is brought to you for free and open access by the Faculty Publications & Research at Poet Commons. It has been accepted for inclusion in Biology by an authorized administrator of Poet Commons. For more information, please contact [email protected]. Parallel changes in genital morphology delineate cryptic diversification of planktonic nudibranchs rspb.royalsocietypublishing.org Celia K. C. Churchill1,†, Alvin Alejandrino2,‡,A´ngel Valde´s2 and Diarmaid O´ Foighil1 1Museum of Zoology, Department of Ecology and Evolutionary Biology, University of Michigan, 1109 Geddes Avenue, Ann Arbor, MI 48109-1079, USA 2Department of Biological Sciences, California State Polytechnic University, 3801 West Temple Avenue, Pomona, Research CA 91768, USA Cite this article: Churchill CKC, Alejandrino A, The relative roles of geographical and non-geographical barriers in the genesis Valde´sA´,O´ Foighil D. 2013 Parallel changes in of genetic isolation are highly debated in evolutionary biology, yet knowing genital morphology delineate cryptic diversifi- how speciation occurs is essential to our understanding of biodiversity. -

Pelagic Sargassum Community Change Over a 40-Year Period: Temporal and Spatial Variability
Mar Biol (2014) 161:2735–2751 DOI 10.1007/s00227-014-2539-y ORIGINAL PAPER Pelagic Sargassum community change over a 40-year period: temporal and spatial variability C. L. Huffard · S. von Thun · A. D. Sherman · K. Sealey · K. L. Smith Jr. Received: 20 May 2014 / Accepted: 3 September 2014 / Published online: 14 September 2014 © The Author(s) 2014. This article is published with open access at Springerlink.com Abstract Pelagic forms of the brown algae (Phaeo- ranging across the Sargasso Sea, Gulf Stream, and south phyceae) Sargassum spp. and their conspicuous rafts are of the subtropical convergence zone. Recent samples also defining characteristics of the Sargasso Sea in the western recorded low coverage by sessile epibionts, both calcifying North Atlantic. Given rising temperatures and acidity in forms and hydroids. The diversity and species composi- the surface ocean, we hypothesized that macrofauna asso- tion of macrofauna communities associated with Sargas- ciated with Sargassum in the Sargasso Sea have changed sum might be inherently unstable. While several biological with respect to species composition, diversity, evenness, and oceanographic factors might have contributed to these and sessile epibiota coverage since studies were con- observations, including a decline in pH, increase in sum- ducted 40 years ago. Sargassum communities were sam- mer temperatures, and changes in the abundance and distri- pled along a transect through the Sargasso Sea in 2011 and bution of Sargassum seaweed in the area, it is not currently 2012 and compared to samples collected in the Sargasso possible to attribute direct causal links. Sea, Gulf Stream, and south of the subtropical conver- gence zone from 1966 to 1975. -

Xoimi AMERICAN COXCIIOLOGY
S31ITnS0NIAN MISCEllANEOUS COLLECTIOXS. BIBLIOGIIAPHY XOimi AMERICAN COXCIIOLOGY TREVIOUS TO THE YEAR 18G0. PREPARED FOR THE SMITHSONIAN INSTITUTION BY . W. G. BINNEY. PART II. FOKEIGN AUTHORS. WASHINGTON: SMITHSONIAN INSTITUTION. JUNE, 1864. : ADYERTISEMENT, The first part of the Bibliography of American Conchology, prepared for the Smithsonian Institution by Mr. Binuey, was published in March, 1863, and embraced the references to de- scriptions of shells by American authors. The second part of the same work is herewith presented to the public, and relates to species of North American shells referred to by European authors. In foreign works binomial authors alone have been quoted, and no species mentioned which is not referred to North America or some specified locality of it. The third part (in an advanced stage of preparation) will in- clude the General Index of Authors, the Index of Generic and Specific names, and a History of American Conchology, together with any additional references belonging to Part I and II, that may be met with. JOSEPH HENRY, Secretary S. I. Washington, June, 1864. (" ) PHILADELPHIA COLLINS, PRINTER. CO]^TENTS. Advertisement ii 4 PART II.—FOREIGN AUTHORS. Titles of Works and Articles published by Foreign Authors . 1 Appendix II to Part I, Section A 271 Appendix III to Part I, Section C 281 287 Appendix IV .......... • Index of Authors in Part II 295 Errata ' 306 (iii ) PART II. FOEEIGN AUTHORS. ( V ) BIBLIOGRxVPHY NOETH AMERICAN CONCHOLOGY. PART II. Pllipps.—A Voyage towards the North Pole, &c. : by CON- STANTiNE John Phipps. Loudou, ITTJc. Pa. BIBLIOGRAPHY OF [part II. FaliricillS.—Fauna Grcenlandica—systematice sistens ani- malia GrcEulandite occidentalis liactenus iudagata, &c., secun dum proprias observatioues Othonis Fabricii. -

A Checklist of Turtle and Whale Barnacles
Journal of the Marine Biological Association of the United Kingdom, 2013, 93(1), 143–182. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S0025315412000847 A checklist of turtle and whale barnacles (Cirripedia: Thoracica: Coronuloidea) ryota hayashi1,2 1International Coastal Research Center, Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5, Kashiwanoha, Kashiwa-shi, Chiba 277-8564 Japan, 2Marine Biology and Ecology Research Program, Extremobiosphere Research Center, Japan Agency for Marine–Earth Science and Technology A checklist of published records of coronuloid barnacles (Cirripedia: Thoracica: Coronuloidea) attached to marine vertebrates is presented, with 44 species (including 15 fossil species) belonging to 14 genera (including 3 fossil genera) and 3 families recorded. Also included is information on their geographical distribution and the hosts with which they occur. Keywords: checklist, turtle barnacles, whale barnacles, Chelonibiidae, Emersoniidae, Coronulidae, Platylepadidae, host and distribution Submitted 10 May 2012; accepted 16 May 2012; first published online 10 August 2012 INTRODUCTION Superorder THORACICA Darwin, 1854 Order SESSILIA Lamarck, 1818 In this paper, a checklist of barnacles of the superfamily Suborder BALANOMORPHA Pilsbry, 1916 Coronuloidea occurring on marine animals is presented. Superfamily CORONULOIDEA Newman & Ross, 1976 The systematic arrangement used herein follows Newman Family CHELONIBIIDAE Pilsbry, 1916 (1996) rather than Ross & Frick (2011) for reasons taken up in Hayashi (2012) in some detail. The present author Genus Chelonibia Leach, 1817 deems the subfamilies of the Cheonibiidae (Chelonibiinae, Chelonibia caretta (Spengler, 1790) Emersoniinae and Protochelonibiinae) proposed by Harzhauser et al. (2011), as well as those included of Ross & Lepas caretta Spengler, 1790: 185, plate 6, figure 5. -

Griffith REVIEW Editon 43: Pacific Highways
Griffith 43 A QUARTERLY OF NEW WRITING & IDEAS GriffithREVIEW43 Pacific Highways ESSAY HINEMOANA BAKER Walking meditations BERNARD BECKETT School report DAVID BURTON A Kiwi feast HAMISH CLAYTON The lie of the land RE KATE DE GOLDI Simply by sailing in a new direction LYNN JENNER Thinking about waves FINLAY MACDONALD Primate city LYNNE McDONALD Cable stations V GREGORY O’BRIEN Patterns of migration ROBERTO ONELL To a neighbour I am getting to know IE ROD ORAM Tectonic Z REBECCA PRIESTLEY Hitching a ride W HARRY RICKETTS On masks and migration JOHN SAKER Born to run CARRIE TIFFANY Reading Geoff Cochrane MATT VANCE An A-frame in Antarctica 43 IAN WEDDE O Salutaris LYDIA WEVERS First, build your hut DAMIEN WILKINS We are all Stan Walker ALISON WONG Pure brightness Highways Pacific ASHLEIGH YOUNG Sea of trees MEMOIR KATE CAMP Whale Road PAMELA ‘JUDY’ ROSS Place in time PETER SWAIN Fitting into the Pacific LEILANI TAMU The beach BRIAN TURNER Open road MoreFREE great eBOOKstories and KATE WOODS Postcard from Beijing poetry are available in PACIFIC HIGHWAYS Vol. 2 REPORTAGE as a free download at SALLY BLUNDELL Amending the map www.griffithreview.com STEVE BRAUNIAS On my way to the border GLENN BUSCH Portrait of an artist FICTION WILLIAM BRANDT Getting to yes EMILY PERKINS Waiheke Island CK STEAD Anxiety POETRY JAMES BROWN GEOFF COCHRANE CLIFF FELL PACIFIC DINAH HAWKEN YA-WEN HO BILL MANHIRE GREGORY O’BRIEN HIGHWAYS VINCENT O’SULLIVAN CO-EDITED BY JULIANNE SCHULTZ ‘Australia’s most stimulating literary journal.’ & LLOYD JONES Cover design: Text Publishing design: Text Cover Canberra Times JOURNAL QUARTERLY Praise for Griffith REVIEW ‘Essential reading for each and every one of us.’ Readings ‘A varied, impressive and international cast of authors.’ The Australian ‘Griffith REVIEW is a must-read for anyone with even a passing interest in current affairs, politics, literature and journalism. -

A New Species of Lepas (Crustacea: Cirripedia: Pedunculata) from the Miocene Mizunami Group, Japan
Bulletin of the Mizunami Fossil Museum, no. 31 (2004), p. 91-93, 1 fig. c 2004, Mizunami Fossil Museum A new species of Lepas (Crustacea: Cirripedia: Pedunculata) from the Miocene Mizunami Group, Japan Hiroaki Karasawa*, Toshio Tanaka* *, and Yoshitsugu Okumura*** * Mizunami Fossil Museum, Yamanouchi, Akeyo, Mizunami, Gifu 509-6132, Japan <[email protected]> ** Aichi Gakuin Junior College, Chikusa, Nagoya 464-8650, Japan < [email protected]> *** Mizunami Fossil Museum, Yamanouchi, Akeyo, Mizunami, Gifu 509-6132, Japan <[email protected]> Abstract Lepas kuwayamai, a new species of a lepadid thracican is described from the lower Miocene Mizunami Group in Gifu Prefecture, Honshu, Japan. This represents the third record for the family from Miocene deposits of Japan. Key words: Crustacea, Cirripedia, Thoracica, Pedunculata, Lepadidae, Lepas, Miocene, Japan Material examined: MFM9043, holotype; 8 paratypes, Introduction MFM9044-9051; Coll. M. Kuwayama in, 2004. All specimens are housed in the Mizunami Fossil Museum. The lepadomorph family Lepadidae Darwin, 1851, is a Diagnosis: Lepas with moderate sized capitulum. Shell small group including six genera (Newman, 1996). Among thick. Scutum triangular, slightly higher than wide, with these, the genus Lepas Linnaeus, 1758, is only known clear growth lines; umbonal tooth absent; very weak from Japan in the fossil record. O’hara et al. (1976) reported radial striae sometimes present; apicoumbonal ridge Lepas sp. from the middle Pleistocene Shimosa Group and weak. Tergum flattened without radial striae; occludent unnamed Miocene species was reported from the lower margin convex, rounded. Carina broad. Miocene Morozaki Group (Mizuno and Takeda, 1993) and Etymology: In honor to our friend, Mr.