MRCK Is Dispensable for Breast Cancer Development in the MMTV
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Profiling Data
Compound Name DiscoveRx Gene Symbol Entrez Gene Percent Compound Symbol Control Concentration (nM) JNK-IN-8 AAK1 AAK1 69 1000 JNK-IN-8 ABL1(E255K)-phosphorylated ABL1 100 1000 JNK-IN-8 ABL1(F317I)-nonphosphorylated ABL1 87 1000 JNK-IN-8 ABL1(F317I)-phosphorylated ABL1 100 1000 JNK-IN-8 ABL1(F317L)-nonphosphorylated ABL1 65 1000 JNK-IN-8 ABL1(F317L)-phosphorylated ABL1 61 1000 JNK-IN-8 ABL1(H396P)-nonphosphorylated ABL1 42 1000 JNK-IN-8 ABL1(H396P)-phosphorylated ABL1 60 1000 JNK-IN-8 ABL1(M351T)-phosphorylated ABL1 81 1000 JNK-IN-8 ABL1(Q252H)-nonphosphorylated ABL1 100 1000 JNK-IN-8 ABL1(Q252H)-phosphorylated ABL1 56 1000 JNK-IN-8 ABL1(T315I)-nonphosphorylated ABL1 100 1000 JNK-IN-8 ABL1(T315I)-phosphorylated ABL1 92 1000 JNK-IN-8 ABL1(Y253F)-phosphorylated ABL1 71 1000 JNK-IN-8 ABL1-nonphosphorylated ABL1 97 1000 JNK-IN-8 ABL1-phosphorylated ABL1 100 1000 JNK-IN-8 ABL2 ABL2 97 1000 JNK-IN-8 ACVR1 ACVR1 100 1000 JNK-IN-8 ACVR1B ACVR1B 88 1000 JNK-IN-8 ACVR2A ACVR2A 100 1000 JNK-IN-8 ACVR2B ACVR2B 100 1000 JNK-IN-8 ACVRL1 ACVRL1 96 1000 JNK-IN-8 ADCK3 CABC1 100 1000 JNK-IN-8 ADCK4 ADCK4 93 1000 JNK-IN-8 AKT1 AKT1 100 1000 JNK-IN-8 AKT2 AKT2 100 1000 JNK-IN-8 AKT3 AKT3 100 1000 JNK-IN-8 ALK ALK 85 1000 JNK-IN-8 AMPK-alpha1 PRKAA1 100 1000 JNK-IN-8 AMPK-alpha2 PRKAA2 84 1000 JNK-IN-8 ANKK1 ANKK1 75 1000 JNK-IN-8 ARK5 NUAK1 100 1000 JNK-IN-8 ASK1 MAP3K5 100 1000 JNK-IN-8 ASK2 MAP3K6 93 1000 JNK-IN-8 AURKA AURKA 100 1000 JNK-IN-8 AURKA AURKA 84 1000 JNK-IN-8 AURKB AURKB 83 1000 JNK-IN-8 AURKB AURKB 96 1000 JNK-IN-8 AURKC AURKC 95 1000 JNK-IN-8 -

Loss of Fam60a, a Sin3a Subunit, Results in Embryonic Lethality and Is Associated with Aberrant Methylation at a Subset of Gene
RESEARCH ARTICLE Loss of Fam60a, a Sin3a subunit, results in embryonic lethality and is associated with aberrant methylation at a subset of gene promoters Ryo Nabeshima1,2, Osamu Nishimura3,4, Takako Maeda1, Natsumi Shimizu2, Takahiro Ide2, Kenta Yashiro1†, Yasuo Sakai1, Chikara Meno1, Mitsutaka Kadota3,4, Hidetaka Shiratori1†, Shigehiro Kuraku3,4*, Hiroshi Hamada1,2* 1Developmental Genetics Group, Graduate School of Frontier Biosciences, Osaka University, Suita, Japan; 2Laboratory for Organismal Patterning, RIKEN Center for Developmental Biology, Kobe, Japan; 3Phyloinformatics Unit, RIKEN Center for Life Science Technologies, Kobe, Japan; 4Laboratory for Phyloinformatics, RIKEN Center for Biosystems Dynamics Research, Kobe, Japan Abstract We have examined the role of Fam60a, a gene highly expressed in embryonic stem cells, in mouse development. Fam60a interacts with components of the Sin3a-Hdac transcriptional corepressor complex, and most Fam60a–/– embryos manifest hypoplasia of visceral organs and die in utero. Fam60a is recruited to the promoter regions of a subset of genes, with the expression of these genes being either up- or down-regulated in Fam60a–/– embryos. The DNA methylation level of the Fam60a target gene Adhfe1 is maintained at embryonic day (E) 7.5 but markedly reduced at –/– *For correspondence: E9.5 in Fam60a embryos, suggesting that DNA demethylation is enhanced in the mutant. [email protected] (SK); Examination of genome-wide DNA methylation identified several differentially methylated regions, [email protected] (HH) which were preferentially hypomethylated, in Fam60a–/– embryos. Our data suggest that Fam60a is †These authors contributed required for proper embryogenesis, at least in part as a result of its regulation of DNA methylation equally to this work at specific gene promoters. -
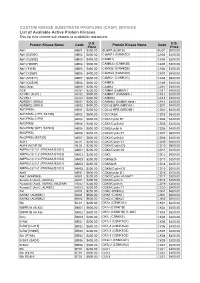
CUSTOM KINASE SUBSTRATE PROFILING (CKSP) SERVICE List of Available Active Protein Kinases This List May Change with Respect to Availability and Pricing
CUSTOM KINASE SUBSTRATE PROFILING (CKSP) SERVICE List of Available Active Protein Kinases This list may change with respect to availability and pricing. Protein Kinase Name Code U.S. Protein Kinase Name Code U.S. Price Price Abl1 AB01 $200.00 BUBR1(BUB1B) BU01 $600.00 Abl1 [E255K] AB02 $400.00 CaMK1δ (CAMK1D) CA03 $200.00 Abl1 [G250E] AB03 $400.00 CAMK1γ CA04 $200.00 Abl1 [H369P] AB04 $600.00 CAMK2α (CAMK2B) CA05 $200.00 Abl1 [T315I] AB05 $400.00 CaMK2β (CAMK2B) CA06 $200.00 Abl1 [Y253F] AB06 $400.00 CaMK2δ (CAMK2D) CA07 $400.00 Abl1 [M351T] AB07 $600.00 CaMK2γ (CAMK2G) CA08 $600.00 Abl1 [Q252H] AB08 $600.00 CAMK3γ CA09 $200.00 Abl2 (Arg) AB09 $200.00 CAMK4 CA10 $200.00 ACK AC01 $200.00 CAMK4 (CaMKIV ) CA11 $400.00 ACVR1 (ALK2 ) AC02 $400.00 CAMKK1 (CAMKKA ) CA12 $200.00 ACVRL1 AC03 $400.00 CAMKK2 CA13 $200.00 ADRBK1 (GRK2) AD01 $200.00 CAMKK2 (CaMKK beta ) CA14 $400.00 ADRBK2 (GRK3) AD02 $400.00 CDC42 BPA (MRCKA ) CD01 $400.00 Akt1/PKBα AK01 $200.00 CDC42 BPB (MRCKB ) CD02 $400.00 Akt1/PKBα [δPH, S473D] AK02 $600.00 CDC7/ASK CD03 $600.00 Akt1/PKBα [δPH] AK03 $600.00 CDK1/cyclin B1 CD04 $400.00 Akt2/PKBβ AK04 $200.00 CDK1/CyclinA2 CD05 $200.00 Akt2/PKBβ [δPH, S474D] AK05 $600.00 CDK2/cyclin A CD06 $400.00 Akt3/PKBγ AK06 $200.00 CDK2/Cyclin E1 CD07 $600.00 Akt3/PKBγ [S472D] AK07 $600.00 CDK2/CyclinA2 CD08 $200.00 ALK1 AL01 $200.00 CDK3/Cyclin E1 CD09 $600.00 ALK4 (ACVR1B) AL02 $200.00 CDK4//Cyclin D3 CD10 $600.00 AMPKα1/β1/γ1 (PRKAA1/B1/G1) AM01 $200.00 CDK4/Cyclin D1 CD11 $200.00 AMPKα1/β1/γ2 (PRKAA1/B1/G2) AM02 $200.00 CDK5 CD12 $600.00 -

Inherited Variation in Mir-290 Expression Suppresses Breast Cancer Progression by Targeting the Metastasis Susceptibility Gene Arid4b
Published OnlineFirst February 27, 2013; DOI: 10.1158/0008-5472.CAN-12-3513 Cancer Tumor and Stem Cell Biology Research Inherited Variation in miR-290 Expression Suppresses Breast Cancer Progression by Targeting the Metastasis Susceptibility Gene Arid4b Natalie Goldberger1, Renard C. Walker1, Chang Hee Kim2, Scott Winter1, and Kent W. Hunter1 Abstract The metastatic cascade is a complex and extremely inefficient process with many potential barriers. Understanding this process is of critical importance because the majority of cancer mortality is associated with metastatic disease. Recently, it has become increasingly clear that microRNAs (miRNA) play important roles in tumorigenesis and metastasis, yet few studies have examined how germline variations may dysregulate miRNAs, in turn affecting metastatic potential. To explore this possibility, the highly metastatic MMTV-PyMT mice were crossed with 25 AKXD (AKR/J Â DBA/2J) recombinant inbred strains to produce F1 progeny with varying metastatic indices. When mammary tumors from the F1 progeny were analyzed by miRNA microarray, miR-290 (containing miR-290-3p and miR-290-5p) was identified as a top candidate progression-associated miRNA. The microarray results were validated in vivo when miR-290 upregulation in two independent breast cancer cell lines suppressed both primary tumor and metastatic growth. Computational analysis identified breast cancer progression gene Arid4b as a top target of miR-290-3p, which was confirmed by luciferase reporter assay. Surprisingly, pathway analysis identified estrogen receptor (ER) signaling as the top canonical pathway affected by miR-290 upregulation. Further analysis showed that ER levels were elevated in miR-290–expressing tumors and positively correlated with apoptosis. -

Mouse Cdc42bpa Knockout Project (CRISPR/Cas9)
https://www.alphaknockout.com Mouse Cdc42bpa Knockout Project (CRISPR/Cas9) Objective: To create a Cdc42bpa knockout Mouse model (C57BL/6J) by CRISPR/Cas-mediated genome engineering. Strategy summary: The Cdc42bpa gene (NCBI Reference Sequence: NM_001033285 ; Ensembl: ENSMUSG00000026490 ) is located on Mouse chromosome 1. 37 exons are identified, with the ATG start codon in exon 1 and the TGA stop codon in exon 37 (Transcript: ENSMUST00000111117). Exon 3~5 will be selected as target site. Cas9 and gRNA will be co-injected into fertilized eggs for KO Mouse production. The pups will be genotyped by PCR followed by sequencing analysis. Note: Exon 3 starts from about 5.22% of the coding region. Exon 3~5 covers 6.33% of the coding region. The size of effective KO region: ~8696 bp. The KO region does not have any other known gene. Page 1 of 10 https://www.alphaknockout.com Overview of the Targeting Strategy Wildtype allele 5' gRNA region gRNA region 3' 1 3 4 5 37 Legends Exon of mouse Cdc42bpa Knockout region Page 2 of 10 https://www.alphaknockout.com Overview of the Dot Plot (up) Window size: 15 bp Forward Reverse Complement Sequence 12 Note: The 2000 bp section upstream of Exon 3 is aligned with itself to determine if there are tandem repeats. No significant tandem repeat is found in the dot plot matrix. So this region is suitable for PCR screening or sequencing analysis. Overview of the Dot Plot (down) Window size: 15 bp Forward Reverse Complement Sequence 12 Note: The 2000 bp section downstream of Exon 5 is aligned with itself to determine if there are tandem repeats. -

Targeting Microcephaly Kinases in Brain Tumors
International Journal of Molecular Sciences Review Precision Revisited: Targeting Microcephaly Kinases in Brain Tumors Gianmarco Pallavicini 1,2,3, Gaia E. Berto 1,2 and Ferdinando Di Cunto 1,2,4,* 1 Neuroscience Institute Cavalieri Ottolenghi, 10126 Turin, Italy; [email protected] (G.P.); [email protected] (G.E.B.) 2 Department of Neurosciences, University of Turin, 10126 Turin, Italy 3 Department of Molecular Biotechnology and Health Sciences, University of Turin, 10126 Turin, Italy 4 Neuroscience Institute of Turin (NIT), 10126 Turin, Italy * Correspondence: [email protected] Received: 26 March 2019; Accepted: 26 April 2019; Published: 28 April 2019 Abstract: Glioblastoma multiforme and medulloblastoma are the most frequent high-grade brain tumors in adults and children, respectively. Standard therapies for these cancers are mainly based on surgical resection, radiotherapy, and chemotherapy. However, intrinsic or acquired resistance to treatment occurs almost invariably in the first case, and side effects are unacceptable in the second. Therefore, the development of new, effective drugs is a very important unmet medical need. A critical requirement for developing such agents is to identify druggable targets required for the proliferation or survival of tumor cells, but not of other cell types. Under this perspective, genes mutated in congenital microcephaly represent interesting candidates. Congenital microcephaly comprises a heterogeneous group of disorders in which brain volume is reduced, in the absence or presence of variable syndromic features. Genetic studies have clarified that most microcephaly genes encode ubiquitous proteins involved in mitosis and in maintenance of genomic stability, but the effects of their inactivation are particularly strong in neural progenitors. -

Molecular Classification of Diffuse Cerebral WHO Grade II/III Gliomas
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2015 Molecular classification of diffuse cerebral WHO grade II/III gliomas using genome- and transcriptome-wide profiling improves stratification of prognostically distinct patient groups Weller, Michael ; Weber, Ruthild G ; Willscher, Edith ; Riehmer, Vera ; Hentschel, Bettina ; Kreuz, Markus ; Felsberg, Jörg ; Beyer, Ulrike ; Löffler-Wirth, Henry ; Kaulich, Kerstin ; Steinbach, Joachim P ; Hartmann, Christian ; Gramatzki, D ; Schramm, J ; Westphal, M ; Schackert, G ; Simon, M ; Martens, T ; Boström, J ; Hagel, C ; Sabel, M ; Krex, D ; Tonn, J C ; Wick, W ; Noell, S ; Schlegel, U ; Radlwimmer, B ; Pietsch, T ; Loeffler, M ; von Deimling, A ; Binder, H ; Reifenberger, G Abstract: Cerebral gliomas of World Health Organization (WHO) grade II and III represent a major challenge in terms of histological classification and clinical management. Here, we asked whether large- scale genomic and transcriptomic profiling improves the definition of prognostically distinct entities. We performed microarray-based genome- and transcriptome-wide analyses of primary tumor samples from a prospective German Glioma Network cohort of 137 patients with cerebral gliomas, including 61 WHO grade II and 76 WHO grade III tumors. Integrative bioinformatic analyses were employed to define molecular subgroups, which were then related to histology, molecular biomarkers, including isocitrate de- hydrogenase 1 or 2 (IDH1/2) mutation, 1p/19q co-deletion and telomerase reverse transcriptase (TERT) promoter mutations, and patient outcome. Genomic profiling identified five distinct glioma groups, in- cluding three IDH1/2 mutant and two IDH1/2 wild-type groups. Expression profiling revealed evidence for eight transcriptionally different groups (five IDH1/2 mutant, three IDH1/2 wild type), which were only partially linked to the genomic groups. -

ARID4B Polyclonal Antibody
For Research Use Only ARID4B Polyclonal antibody Catalog Number:24499-1-AP 3 Publications www.ptgcn.com Catalog Number: GenBank Accession Number: Recommended Dilutions: Basic Information 24499-1-AP BC130418 WB 1:500-1:1000 Size: GeneID (NCBI): IP 0.5-4.0 ug for IP and 1:500-1:2000 1000 μg/ml 51742 for WB IHC 1:20-1:200 Source: Full Name: IF 1:10-1:100 Rabbit AT rich interactive domain 4B (RBP1- Isotype: like) IgG Calculated MW: Purification Method: 1312 aa, 148 kDa Antigen affinity purification Observed MW: Immunogen Catalog Number: 100 kDa, 200 kDa AG21462 Applications Tested Applications: Positive Controls: IF, IHC, IP, WB,ELISA WB : MCF-7 cells; A549 cells, K-562 cells Cited Applications: IP : MCF-7 cells; WB IHC : human testis tissue; human breast cancer tissue Species Specificity: human IF : MCF-7 cells; Cited Species: human Note-IHC: suggested antigen retrieval with TE buffer pH 9.0; (*) Alternatively, antigen retrieval may be performed with citrate buffer pH 6.0 ARID4B, also named as BRCAA1 or SAP180, is a 1312 amino acid protein, which contains one ARID domain. ARID4B Background Information localizes in the nucleus and cytoplasm. ARID4B as a transcription repressor may function in the assembly and/or enzymatic activity of the Sin3A corepressor complex or in mediating interactions between the complex and other regulatory complexes. ARID4B is highly expressed in the testis and in breast, lung, colon, pancreatic and ovarian cancers. Expressed at low levels in the thymus, prostate and ovary. Notable Publications Author Pubmed ID Journal Application Ti-Chun Chan 33052246 Theranostics WB Xiangxiang Liu 30305607 Cell Death Dis WB Wanjin Li 28805661 J Clin Invest WB Storage: Storage Store at -20ºC. -

Activation of Diverse Signalling Pathways by Oncogenic PIK3CA Mutations
ARTICLE Received 14 Feb 2014 | Accepted 12 Aug 2014 | Published 23 Sep 2014 DOI: 10.1038/ncomms5961 Activation of diverse signalling pathways by oncogenic PIK3CA mutations Xinyan Wu1, Santosh Renuse2,3, Nandini A. Sahasrabuddhe2,4, Muhammad Saddiq Zahari1, Raghothama Chaerkady1, Min-Sik Kim1, Raja S. Nirujogi2, Morassa Mohseni1, Praveen Kumar2,4, Rajesh Raju2, Jun Zhong1, Jian Yang5, Johnathan Neiswinger6, Jun-Seop Jeong6, Robert Newman6, Maureen A. Powers7, Babu Lal Somani2, Edward Gabrielson8, Saraswati Sukumar9, Vered Stearns9, Jiang Qian10, Heng Zhu6, Bert Vogelstein5, Ben Ho Park9 & Akhilesh Pandey1,8,9 The PIK3CA gene is frequently mutated in human cancers. Here we carry out a SILAC-based quantitative phosphoproteomic analysis using isogenic knockin cell lines containing ‘driver’ oncogenic mutations of PIK3CA to dissect the signalling mechanisms responsible for oncogenic phenotypes induced by mutant PIK3CA. From 8,075 unique phosphopeptides identified, we observe that aberrant activation of PI3K pathway leads to increased phosphorylation of a surprisingly wide variety of kinases and downstream signalling networks. Here, by integrating phosphoproteomic data with human protein microarray-based AKT1 kinase assays, we discover and validate six novel AKT1 substrates, including cortactin. Through mutagenesis studies, we demonstrate that phosphorylation of cortactin by AKT1 is important for mutant PI3K-enhanced cell migration and invasion. Our study describes a quantitative and global approach for identifying mutation-specific signalling events and for discovering novel signalling molecules as readouts of pathway activation or potential therapeutic targets. 1 McKusick-Nathans Institute of Genetic Medicine and Department of Biological Chemistry, Johns Hopkins University School of Medicine, 733 North Broadway, BRB 527, Baltimore, Maryland 21205, USA. -

Anti-CDC42BPA Polyclonal Antibody (CABT- BL1018) This Product Is for Research Use Only and Is Not Intended for Diagnostic Use
Anti-CDC42BPA polyclonal antibody (CABT- BL1018) This product is for research use only and is not intended for diagnostic use. PRODUCT INFORMATION Immunogen Peptide corresponding to the amino acids 1671-1685 of human MRCK alpha Isotype IgG Source/Host Rabbit Species Reactivity Human Conjugate Unconjugated Applications WB, ICC Buffer The reagent is provided in phosphate buffered saline (PBS) containing26mM sodium azide and 0.2% (w/v) high-grade protease free Bovine Serum Albumin (BSA) as a stabilizing agent. Preservative 26mM Sodium Azide Storage Store at 2-8°C. Do not use after expiration date stamped on vial label. For long-term storage aliquot and store at -20°C. Avoid freeze/thaw cycles. BACKGROUND Introduction The protein encoded by this gene is a member of the Serine/Threonine protein kinase family. This kinase contains multiple functional domains. Its kinase domain is highly similar to that of the myotonic dystrophy protein kinase (DMPK). This kinase also contains a Rac interactive binding (CRIB) domain, and has been shown to bind CDC42. It may function as a CDC42 downstream effector mediating CDC42 induced peripheral actin formation, and promoting cytoskeletal reorganization. Multiple alternatively spliced transcript variants have been described, and the full- length nature of two of them has been reported. [provided by RefSeq, Jul 2008] GENE INFORMATION 45-1 Ramsey Road, Shirley, NY 11967, USA Email: [email protected] Tel: 1-631-624-4882 Fax: 1-631-938-8221 1 © Creative Diagnostics All Rights Reserved Entrez Gene ID 8476 Protein Refseq NP_003598 UniProt ID Q5VT25 45-1 Ramsey Road, Shirley, NY 11967, USA Email: [email protected] Tel: 1-631-624-4882 Fax: 1-631-938-8221 2 © Creative Diagnostics All Rights Reserved. -

PRODUCTS and SERVICES Target List
PRODUCTS AND SERVICES Target list Kinase Products P.1-11 Kinase Products Biochemical Assays P.12 "QuickScout Screening Assist™ Kits" Kinase Protein Assay Kits P.13 "QuickScout Custom Profiling & Panel Profiling Series" Targets P.14 "QuickScout Custom Profiling Series" Preincubation Targets Cell-Based Assays P.15 NanoBRET™ TE Intracellular Kinase Cell-Based Assay Service Targets P.16 Tyrosine Kinase Ba/F3 Cell-Based Assay Service Targets P.17 Kinase HEK293 Cell-Based Assay Service ~ClariCELL™ ~ Targets P.18 Detection of Protein-Protein Interactions ~ProbeX™~ Stable Cell Lines Crystallization Services P.19 FastLane™ Structures ~Premium~ P.20-21 FastLane™ Structures ~Standard~ Kinase Products For details of products, please see "PRODUCTS AND SERVICES" on page 1~3. Tyrosine Kinases Note: Please contact us for availability or further information. Information may be changed without notice. Expression Protein Kinase Tag Carna Product Name Catalog No. Construct Sequence Accession Number Tag Location System HIS ABL(ABL1) 08-001 Full-length 2-1130 NP_005148.2 N-terminal His Insect (sf21) ABL(ABL1) BTN BTN-ABL(ABL1) 08-401-20N Full-length 2-1130 NP_005148.2 N-terminal DYKDDDDK Insect (sf21) ABL(ABL1) [E255K] HIS ABL(ABL1)[E255K] 08-094 Full-length 2-1130 NP_005148.2 N-terminal His Insect (sf21) HIS ABL(ABL1)[T315I] 08-093 Full-length 2-1130 NP_005148.2 N-terminal His Insect (sf21) ABL(ABL1) [T315I] BTN BTN-ABL(ABL1)[T315I] 08-493-20N Full-length 2-1130 NP_005148.2 N-terminal DYKDDDDK Insect (sf21) ACK(TNK2) GST ACK(TNK2) 08-196 Catalytic domain -

Supplementary Table 5.List of the 220 Most Frequently Amplified Genes In
Supplementary Table 5. List of the 220 most frequently amplified genes in this study. The table includes their chromosomal location, the amplification frequency in ER-positive female breast cancer with associated p-value for difference in proportions, the preference for surrogate intrinsic molecular subtype, and associations with clinical, pathological and genetic characteristics. Potentially druggable gene categories, clinical actionability and known drug interactions are indicated per gene. Gene Full name chr location % amp in FFPE % amps in FF total % amp % amp ER+ FBC* p-value MBC vs ER+ FBC** % in lumA-like % in lumB-like p-value BRCA2 germline Age Hist type ER status PR status HER2 status Grade MAI Size LN SNV load PIK3CA mut KM (OS)*** KM (5Y OS)*** druggable gene category# clinically actionable?## known drug interactions?### THBS1 thrombospondin 1 15q14 37% 9% 30% 0.1% <0.0001 23% 35% 0.128 ns ns ns ns ns ns ns ns ns ns ns ns 0.642 p=0.832 cell surface, tumor suppressor, drug resistance, external side of plasma membrane no none PRKDC protein kinase, DNA-activated, catalytic polypeptide 8q11.21 35% 7% 27% 10.9% <0.0001 26% 30% 0.595 ns ns ns ns ns ns ns ns ns ns ns ns 0.838 p=0.903 (serine threonine) kinase, druggable genome, PI3 kinase, tumor suppressor, TF complex, TF binding, DNA repair yes DNA-PK INHIBITOR V (DNA-PK inhibitor); WORTMANNIN (PI3K inhibitor); SF1126 (PI3 kinase/mTOR inhibitor) TBX3 T-box 3 12q24.21 34% 7% 27% 0.1% <0.0001 20% 35% 0.053 ns ns ns ns ns ns ns ns ns ns ns ns 0.439 p=0.264 tumor suppressor, TF binding