Proquest Dissertations
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sherwood Forest Lepidoptera Species List
Sherwood Forest Lepidoptera Historical and modern species lists and records - First Edition 2009 T. and D. Pendleton Introduction to Sherwood Forest Lepidoptera First edition 2009 One aspect of recording which has not changed since the 1800’s, is the lack of attention given by Lepidopterists to the site’s micro lepidoptera. Virtually all This publication is aimed at providing an accurate record of all Sherwood Forest micro records sourced from Carr’s book were attributed to R.E. Brameld, who Lepidoptera recorded from the 1800‘s, the 1900’s, post-2000 records and must have spent many hours in the field recording. Without his work, then much completely up to date with the inclusion of the latest records from 2008 and larger gaps would have appeared in the records. As it is, much work is still follows the same format as we have used in the accompanying publication required to record the many leaf-mining species which Sherwood Forest Sherwood Forest Coleoptera - 2009 Edition. undoubtedly holds. It is by no means a complete list and should not be treated as so, but Recent Lepidoptera surveys and recording represents a hopeful start on the way to producing a definitive Sherwood Forest site list, by trying to gather all available Lepidoptera records in one document. A great deal of moth trapping and recording took place at the Visitor Centre during the 1980’s and has provided some very useful data, which at some time This is the first edition of this project and includes traceable public records in the future, will perhaps be used to enable a comparison with more recent and those held privately by local naturalists and our thanks go to those records and numbers. -

Monitoring Report Spring/Summer 2015 Contents
Wimbledon and Putney Commons Monitoring Report Spring/Summer 2015 Contents CONTEXT 1 A. SYSTEMATIC RECORDING 3 METHODS 3 OUTCOMES 6 REFLECTIONS AND RECOMMENDATIONS 18 B. BIOBLITZ 19 REFLECTIONS AND LESSONS LEARNT 21 C. REFERENCES 22 LIST OF FIGURES Figure 1 Location of The Plain on Wimbledon and Putney Commons 2 Figure 2 Experimental Reptile Refuge near the Junction of Centre Path and Somerset Ride 5 Figure 3 Contrasting Cut and Uncut Areas in the Conservation Zone of The Plain, Spring 2015 6/7 Figure 4 Notable Plant Species Recorded on The Plain, Summer 2015 8 Figure 5 Meadow Brown and white Admiral Butterflies 14 Figure 6 Hairy Dragonfly and Willow Emerald Damselfly 14 Figure 7 The BioBlitz Route 15 Figure 8 Vestal and European Corn-borer moths 16 LIST OF TABLES Table 1 Mowing Dates for the Conservation Area of The Plain 3 Table 2 Dates for General Observational Records of The Plain, 2015 10 Table 3 Birds of The Plain, Spring - Summer 2015 11 Table 4 Summary of Insect Recording in 2015 12/13 Table 5 Rare Beetles Living in the Vicinity of The Plain 15 LIST OF APPENDICES A1 The Wildlife and Conservation Forum and Volunteer Recorders 23 A2 Sward Height Data Spring 2015 24 A3 Floral Records for The Plain : Wimbledon and Putney Commons 2015 26 A4 The Plain Spring and Summer 2015 – John Weir’s General Reports 30 A5 a Birds on The Plain March to September 2015; 41 B Birds on The Plain - summary of frequencies 42 A6 ai Butterflies on The Plain (DW) 43 aii Butterfly long-term transect including The Plain (SR) 44 aiii New woodland butterfly transect -

Desktop Biodiversity Report
Desktop Biodiversity Report Lindfield Rural and Urban Parishes ESD/14/65 Prepared for Terry Oliver 10th February 2014 This report is not to be passed on to third parties without prior permission of the Sussex Biodiversity Record Centre. Please be aware that printing maps from this report requires an appropriate OS licence. Sussex Biodiversity Record Centre report regarding land at Lindfield Rural and Urban Parishes 10/02/2014 Prepared for Terry Oliver ESD/14/65 The following information is enclosed within this report: Maps Sussex Protected Species Register Sussex Bat Inventory Sussex Bird Inventory UK BAP Species Inventory Sussex Rare Species Inventory Sussex Invasive Alien Species Full Species List Environmental Survey Directory SNCI L61 - Waspbourne Wood; M08 - Costells, Henfield & Nashgill Woods; M10 - Scaynes Hill Common; M18 - Walstead Cemetery; M25 - Scrase Valley Local Nature Reserve; M49 - Wickham Woods. SSSI Chailey Common. Other Designations/Ownership Area of Outstanding Natural Beauty; Environmental Stewardship Agreement; Local Nature Reserve; Notable Road Verge; Woodland Trust Site. Habitats Ancient tree; Ancient woodland; Coastal and floodplain grazing marsh; Ghyll woodland; Traditional orchard. Important information regarding this report It must not be assumed that this report contains the definitive species information for the site concerned. The species data held by the Sussex Biodiversity Record Centre (SxBRC) is collated from the biological recording community in Sussex. However, there are many areas of Sussex where the records held are limited, either spatially or taxonomically. A desktop biodiversity report from the SxBRC will give the user a clear indication of what biological recording has taken place within the area of their enquiry. -
North American Ypsolopha (Ypsolophidae): Losing Some And
Revision of North American Ypsolopha with new taxa and synonyms Jae-Cheon Sohn Department of Science Education, Gongju National University of Education, Chungnam, Republic of Korea Introduction Ypsolopha is the type genus of Ypsolophidae sensu Kyrki (1990) which is sometimes treated as a distinct group of Plutellidae. Ypsolopha has more than 120 Y. aleutianella Y. angelicella Y. arizonella Y. barberella described species worldwide, with the greatest diversity in North America and East Asia; 34 species have been recorded from the Nearctic (Walsingham, 1881, 1889; Beutenmüller, 1889; Busck, 1903, 1906; Kearfott, 1907; Y. barberella (form flavistrigella) Y. cervella Y. cervella (form subsylvella) Y. cockerella Braun, 1925; Meyrick, 1927; Powell, 1967; Heppner, 1982). However, the taxonomic resolution of Ypsolopha in the North America still remains incomplete. Our revision of the North American Ypsolopha revealed nine new species, six new synonyms and one resurrected Y. delicatella Y. dentella Y. dorsimaculella Y. falciferella species. As a result, we suggest a new estimate of Ypsolopha species diversity in North America to consist of more than 37 species. Y. gerdanella Y. leptaula Y. lyonothymnae Y. nella Ypsolopha Latreille, 1796 Y. querciella Y. rubrella Y. schwarziella Y. senex Laterial view of head of Y. laorum (after Robinson et al., 1994) Y. striatella Y. sublucella Y. undulatella Y. ustella Resting posture of Y. blandella Y. vinterella Y. walsinghamiella 2) New Species of Ypsolopha from North America A total of nine new species have been recognized (eight of them shown here). Male and female of Y. contractella, Wing venation of Y. vittella (modified from Moriuti, 1977) Materials & Methods n. -

1 of 9 the Hidden Gardens Species List 17/01/2018 Group Taxon
1 of 9 The Hidden Gardens 17/01/2018 species list Group Taxon Common Name Earliest Latest Records acarine Acari various mites 2017 2017 4 annelid Lumbricus rubellus Red Worm 2017 2017 1 annelid Lumbricus terrestris Common Earthworm 2017 2017 1 bird Accipiter nisus Sparrowhawk 2016 2016 1 bird Aegithalos caudatus Long-tailed Tit 2012 2017 4 bird Carduelis carduelis Goldfinch 2012 2016 3 bird Chloris chloris Greenfinch 2012 2017 4 bird Columba livia Feral Pigeon 2012 2017 3 bird Columba palumbus Woodpigeon 2013 2017 2 bird Corvus corone Carrion Crow 2016 2016 1 bird Corvus monedula Jackdaw 2017 2017 1 bird Cyanistes caeruleus Blue Tit 2012 2017 4 bird Erithacus rubecula Robin 2012 2017 4 bird Fringilla coelebs Chaffinch 2012 2017 2 bird Larus canus Common Gull 2012 2012 1 bird Larus fuscus Lesser Black-backed Gull 2015 2015 2 bird Motacilla alba Pied Wagtail 2017 2017 1 bird Parus major Great Tit 2012 2017 3 bird Passer domesticus House Sparrow 2012 2012 1 bird Periparus ater Coal Tit 2013 2015 2 bird Phylloscopus collybita Chiffchaff 2014 2014 1 bird Phylloscopus trochilus Willow Warbler 2013 2015 2 bird Pica pica Magpie 2012 2017 3 bird Prunella modularis Dunnock 2012 2017 3 bird Pyrrhula pyrrhula Bullfinch 2012 2017 4 bird Regulus regulus Goldcrest 2016 2017 3 bird Spinus spinus Siskin 2012 2012 1 bird Streptopelia decaocto Collared Dove 2012 2012 1 bird Sturnus vulgaris Starling 2013 2015 2 bird Sylvia atricapilla Blackcap 2015 2015 2 bird Troglodytes troglodytes Wren 2012 2017 3 bird Turdus iliacus Redwing 2016 2016 2 bird Turdus merula -

Western Palaearctic Oedicephalini and Phaeogenini
2014, Entomologist’s Gazette 65: 109–129 Western Palaearctic Oedicephalini and Phaeogenini (Hymenoptera: Ichneumonidae, Ichneumoninae) in the National Museums of Scotland, with distributional data including 28 species new to Britain, rearing records, and descriptions of two new species of Aethecerus Wesmael and one of Diadromus Wesmael ERICH DILLER Zoologische Staatssammulung München, Münchhausenstrasse 21, D–81247 München, Germany [email protected] MARK R. SHAW1 National Museums of Scotland, Chambers Street, Edinburgh EH1 1JF, U.K. [email protected] Synopsis An account is given of approximately 3,250 western Palaearctic specimens, comprising 110 determined species, of the tribes Oedicephalini and Phaeogenini in the National Museums of Scotland. Distributional and phenological data are given for all species, and rearing records are provided for about 50, although not always with the host’s identity fully clear. Twenty eight species are newly recorded from Britain, of which Aethecerus horstmanni sp. nov., Aethecerus ruberpedatus sp. nov. and Diadromus nitidigaster sp. nov. are described and figured. Tycherus histrio (Wesmael, 1848) sp. rev. is raised from synonymy with Tycherus ischiomelinus (Gravenhorst, 1829); Dicaelotus schmiedeknechti nom. nov. is provided for Dicaelotus ruficornis (Schmiedeknecht, 1903) nec (Ashmead, 1890); Aethecerus subuliferus (Holmgren, 1890) is proposed as a comb. nov.; and Phaeogenes nigridens Wesmael, 1848, is proposed as a comb. rev. Keywords: Ichneumonidae, Ichneumoninae, Oedicephalini, Phaeogenini, parasitoids, taxonomy, phenology, distribution, hosts, Lepidoptera, British Isles. Introduction The National Museums of Scotland (NMS) has an extensive collection of western Palaearctic Ichneumonoidea which is mostly of fairly recent origin (since about 1980) and has a reasonably good standard of specimen preparation and data. -

Harmful Non-Indigenous Species in the U.S
Harmful Non-Indigenous Species in the United States September 1993 OTA-F-565 NTIS order #PB94-107679 GPO stock #052-003-01347-9 Recommended Citation: U.S. Congress, Office of Technology Assessment, Harmful Non-Indigenous Species in the United States, OTA-F-565 (Washington, DC: U.S. Government Printing Office, September 1993). For Sale by the U.S. Government Printing Office ii Superintendent of Documents, Mail Stop, SSOP. Washington, DC 20402-9328 ISBN O-1 6-042075-X Foreword on-indigenous species (NIS)-----those species found beyond their natural ranges—are part and parcel of the U.S. landscape. Many are highly beneficial. Almost all U.S. crops and domesticated animals, many sport fish and aquiculture species, numerous horticultural plants, and most biologicalN control organisms have origins outside the country. A large number of NIS, however, cause significant economic, environmental, and health damage. These harmful species are the focus of this study. The total number of harmful NIS and their cumulative impacts are creating a growing burden for the country. We cannot completely stop the tide of new harmful introductions. Perfect screening, detection, and control are technically impossible and will remain so for the foreseeable future. Nevertheless, the Federal and State policies designed to protect us from the worst species are not safeguarding our national interests in important areas. These conclusions have a number of policy implications. First, the Nation has no real national policy on harmful introductions; the current system is piecemeal, lacking adequate rigor and comprehensiveness. Second, many Federal and State statutes, regulations, and programs are not keeping pace with new and spreading non-indigenous pests. -

BIODIVERSITY and ENVIRONMENT of NEW ROAD, LITTLE LONDON and NEIGHBOURING COUNTRYSIDE by Dr Paul Sterry Contents: 1
BIODIVERSITY AND ENVIRONMENT OF NEW ROAD, LITTLE LONDON AND NEIGHBOURING COUNTRYSIDE by Dr Paul Sterry Contents: 1. Summary. 2. A brief history. 3. Notable habitats alongside New Road and in the neighbouring countryside. 4. Protected and notable species found on New Road and in the surrounding countryside. Appendix 1 - Historical land use in Little London and its influence on biodiversity. Appendix 2 - Lepidoptera (Butterflies and Moths) recorded on New Road, Little London 2004-2019 (generalised OS Grid Reference SU6159). Appendix 3 - Ageing Hedgerows. About the author : Paul Sterry has BSc and PhD in Zoology and Ecology from Imperial College, London. After 5 years as a Research Fellow at the University of Sussex working on freshwater ecology he embarked on a freelance career as a wildlife author and photographer. Over the last 35 years he has written and illustrated more than 50 books, concentrating mainly on British Wildlife, with the emphasis on photographic field guides. Best-selling titles include Collins Complete British Trees, Collins Complete British Wildlife and Collins Life-size Birds. Above: Barn Owl flying over grassland in the neighbourhood of New Road. 1. Summary Located in the Parish of Pamber, Little London is a Biodiversity hotspot with New Road at its environmental heart. Despite the name New Road is one of the oldest highways in the village and this is reflected in the range of wildlife found along its length, and in the countryside bordering it. New Road has significance for wildlife far beyond is narrow, single-track status. Its ancient hedgerows and adjacent meadows are rich in wildlife but of equal importance is its role as a corridor of wildlife connectivity. -

The Lepidoptera Families and Associated Orders of British Columbia
The Lepidoptera Families and Associated Orders of British Columbia The Lepidoptera Families and Associated Orders of British Columbia G.G.E. Scudder and R.A. Cannings March 31, 2007 G.G.E. Scudder and R.A. Cannings Printed 04/25/07 The Lepidoptera Families and Associated Orders of British Columbia 1 Table of Contents Introduction ................................................................................................................................5 Order MEGALOPTERA (Dobsonflies and Alderflies) (Figs. 1 & 2)...........................................6 Description of Families of MEGALOPTERA .............................................................................6 Family Corydalidae (Dobsonflies or Fishflies) (Fig. 1)................................................................6 Family Sialidae (Alderflies) (Fig. 2)............................................................................................7 Order RAPHIDIOPTERA (Snakeflies) (Figs. 3 & 4) ..................................................................9 Description of Families of RAPHIDIOPTERA ...........................................................................9 Family Inocelliidae (Inocelliid snakeflies) (Fig. 3) ......................................................................9 Family Raphidiidae (Raphidiid snakeflies) (Fig. 4) ...................................................................10 Order NEUROPTERA (Lacewings and Ant-lions) (Figs. 5-16).................................................11 Description of Families of NEUROPTERA ..............................................................................12 -
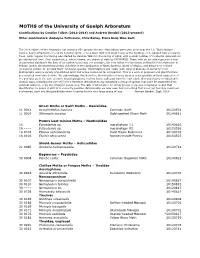
Moth Records
MOTHS of the University of Guelph Arboretum Contributions by Candice Talbot (2012-2014) and Andrew Bendall (2013-present) Other contributors: Andalyne Tofflemire, Chris Earley, Fiona Reid, Mike Kent The 2018 edition of the Arboretum list contains 851 species of moth. Most adults were seen at or near the J.C. Taylor Nature Centre, being attracted to the white building lights, or to a black light and sheet hung by the building, or to painted bait on nearby trees. Semi-regular monitoring was started by Candice Talbot in the spring of 2012, with a small number of incidental observations pre-dating that time. First record dates, where known, are shown at right as YYYYMMDD. Those with an asterisk represent a first documented sighting if the date of an earlier record was not available. We now follow the taxonomy outlined in Pohl, Patterson & Pelham (2016) Annotated taxonomic checklist of the Lepidoptera of North America, North of Mexico, and adopt their revised numbering system for all valid North American species. Identifications are made (with varying degrees of certainty) from photographs using a variety of published print and online resources for comparison. This is a work in progress and identifications are revisited from time to time. We acknowledge that definitive identification of some species is not possible without dissection of the genitalia or, in the case of some microlepidoptera, rearing larvae collected from the host plant. Our uncertainty is indicated in various ways, including the use of [t] for a tentative identification, by indicating a group of species that can't be separated from external features, or by identifying to genus only. -

Identification of Micro-Moth Families
Identification of micro-moth families Introduction This guide is intended for use by Garden Moth Scheme (GMS) recorders and other British and Irish moth enthusiasts. It is aimed at those who already have some understanding of moth taxonomy and morphology but struggle to easily home-in on which moth family a micro- moth belongs to. For further information about how to start out moth- trapping see www.gardenmoths.org.uk. There are currently over 1600 British and Irish micro-moth species, compared to around 800 species of macro-moth and 60 species of butterfly, so species identification is particularly challenging. Narrowing down identification to species requires additional information from training courses, websites and field guides. This guide shows photographs of 45 micro-moth families at rest, together with identification tips, similar species, and the number of species in that family. Taxonomists disagree on the order and arrangement of families, but this guide follows the numbering system of Bradley and Fletcher (B&F) wherever possible. The guide is intended to be used with the excellent Field Guide to the Micromoths of Great Britain and Ireland by Sterling, Parsons and Lewington, British Wildlife Publishing (2012, SPL), which covers 1000+ micro-moth species. However, for the uninitiated, facing a light trap full of micro-moths starting from scratch with SPL can be both daunting and time-consuming. Family names are different and are shown in a different order in SPL and B&F, so in order to reduce confusion, after the B&F numbers are quoted for each family the page numbers for the plates in SPL are also given. -

Folkestone and Hythe Birds Tetrad Guide: TR23 P (Capel-Le-Ferne and Folkestone Warren East)
Folkestone and Hythe Birds Tetrad Guide: TR23 P (Capel-le-Ferne and Folkestone Warren East) The cliff-top provides an excellent vantage point for monitoring visual migration and has been well-watched by Dale Gibson, Ian Roberts and others since 1991. The first promontory to the east of the Cliff-top Café is easily accessible and affords fantastic views along the cliffs and over the Warren. The elevated postion can enable eye-level views of arriving raptors which often use air currents over the Warren to gain height before continuing inland. A Rough-legged Buzzard, three Black Kites, three Montagu’s Harriers and numerous Honey Buzzards, Red Kites, Marsh Harriers and Ospreys have been recorded. It is also perfect for watching arriving swifts and hirundines which may pause to feed over the Warren. Alpine Swift has occurred on three occasions and no less than nine Red- rumped Swallows have been logged here. Looking west from near the Cliff-top Café towards Copt Point and Folkestone Looking east from near the Cliff-top Café towards Abbotscliffe The Warren below the Cliff-top Café Looking west from the bottom of the zigzag path Visual passage will also comprise Sky Larks, Starlings, thrushes, wagtails, pipits, finches and buntings in season, whilst scarcities have included Tawny Pipit, Golden Oriole, Serin, Hawfinch and Snow Bunting. Other oddities have included Purple Heron, Short-eared Owl, Little Ringed Plover, Ruff and Ring-necked Parakeet, whilst in June 1992 a Common Rosefinch was seen on the cliff edge. Below the Cliff-top Café there is a zigzag path leading down into the Warren.