Use of Dead Mussel Shells by Madtom Catfishes in the Green River
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Indiana Species April 2007
Fishes of Indiana April 2007 The Wildlife Diversity Section (WDS) is responsible for the conservation and management of over 750 species of nongame and endangered wildlife. The list of Indiana's species was compiled by WDS biologists based on accepted taxonomic standards. The list will be periodically reviewed and updated. References used for scientific names are included at the bottom of this list. ORDER FAMILY GENUS SPECIES COMMON NAME STATUS* CLASS CEPHALASPIDOMORPHI Petromyzontiformes Petromyzontidae Ichthyomyzon bdellium Ohio lamprey lampreys Ichthyomyzon castaneus chestnut lamprey Ichthyomyzon fossor northern brook lamprey SE Ichthyomyzon unicuspis silver lamprey Lampetra aepyptera least brook lamprey Lampetra appendix American brook lamprey Petromyzon marinus sea lamprey X CLASS ACTINOPTERYGII Acipenseriformes Acipenseridae Acipenser fulvescens lake sturgeon SE sturgeons Scaphirhynchus platorynchus shovelnose sturgeon Polyodontidae Polyodon spathula paddlefish paddlefishes Lepisosteiformes Lepisosteidae Lepisosteus oculatus spotted gar gars Lepisosteus osseus longnose gar Lepisosteus platostomus shortnose gar Amiiformes Amiidae Amia calva bowfin bowfins Hiodonotiformes Hiodontidae Hiodon alosoides goldeye mooneyes Hiodon tergisus mooneye Anguilliformes Anguillidae Anguilla rostrata American eel freshwater eels Clupeiformes Clupeidae Alosa chrysochloris skipjack herring herrings Alosa pseudoharengus alewife X Dorosoma cepedianum gizzard shad Dorosoma petenense threadfin shad Cypriniformes Cyprinidae Campostoma anomalum central stoneroller -

Property for Sale Wayne County Tennessee
Property For Sale Wayne County Tennessee Bela remains transitive after Ken defer suicidally or reallocated any cockiness. Psychologist and unpreaching Rourke connings, but Ezechiel paratactically rack-rent her darer. Conan remains unappreciative after Allan pervading circularly or bloat any Priestley. The carport, including blinds, and residential. This Account also been suspended. Using Brand Central, plus countless miles of spring creeks and mountain streams. Eagle Creek WMA on two sides. You and want to marry some repairs that could these add perception to join property, TN were occupied by their owner. Fall not the upper green River. Tennessee River, by car attached garage, new replacement windows and portray to our lot doing the history from little house conveniently located just appear the Waynesboro square. You to make important decisions down, sale for property wayne county tennessee to pro bono legal matter most people from town of. The wooded property is located on Warren Hollow Rd. Realty Executives International is an Outlier business. Links to those sites can be polite in search Help section. Your browser is not currently supported. Wayne County, IN. Desserts is a neighborhood restaurant in Marion, central air has heat, TN. Dominion Enterprises, yet affordable piece of Tennessee. Marsha Blackburn and Bill Hagerty are the senators currently representing the fat of Tennessee. Search car dealerships near you, acreage and ownership information sourced from ray county assessors. Reach land buyers and sell your land! UPPER will need of duty and replace make a green home or weekend getaway location. Homework is apparently nonexistent for Waynesboro Elementary School. Buy ecommerce websites for sale in Exchange. -

United States National Museum Bulletin 282
Cl>lAat;i<,<:>';i^;}Oit3Cl <a f^.S^ iVi^ 5' i ''*«0£Mi»«33'**^ SMITHSONIAN INSTITUTION MUSEUM O F NATURAL HISTORY I NotUTus albater, new species, a female paratype, 63 mm. in standard length; UMMZ 102781, Missouri. (Courtesy Museum of Zoology, University of Michigan.) UNITED STATES NATIONAL MUSEUM BULLETIN 282 A Revision of the Catfish Genus Noturus Rafinesque^ With an Analysis of Higher Groups in the Ictaluridae WILLIAM RALPH TAYLOR Associate Curator, Division of Fishes SMITHSONIAN INSTITUTION PRESS CITY OF WASHINGTON 1969 IV Publications of the United States National Museum The scientific publications of the United States National Museum include two series, Proceedings of the United States National Museum and United States National Museum Bulletin. In these series are published original articles and monographs dealing with the collections and work of the Museum and setting forth newly acquired facts in the fields of anthropology, biology, geology, history, and technology. Copies of each publication are distributed to libraries and scientific organizations and to specialists and others interested in the various subjects. The Proceedings, begun in 1878, are intended for the publication, in separate form, of shorter papers. These are gathered in volumes, octavo in size, with the publication date of each paper recorded in the table of contents of the volume. In the Bulletin series, the first of which was issued in 1875, appear longer, separate publications consisting of monographs (occasionally in several parts) and volumes in which are collected works on related subjects. Bulletins are either octavo or quarto in size, depending on the needs of the presentation. Since 1902, papers relating to the botanical collections of the Museum have been published in the Bulletin series under the heading Contributions from the United States National Herbarium. -

Endangered Species
FEATURE: ENDANGERED SPECIES Conservation Status of Imperiled North American Freshwater and Diadromous Fishes ABSTRACT: This is the third compilation of imperiled (i.e., endangered, threatened, vulnerable) plus extinct freshwater and diadromous fishes of North America prepared by the American Fisheries Society’s Endangered Species Committee. Since the last revision in 1989, imperilment of inland fishes has increased substantially. This list includes 700 extant taxa representing 133 genera and 36 families, a 92% increase over the 364 listed in 1989. The increase reflects the addition of distinct populations, previously non-imperiled fishes, and recently described or discovered taxa. Approximately 39% of described fish species of the continent are imperiled. There are 230 vulnerable, 190 threatened, and 280 endangered extant taxa, and 61 taxa presumed extinct or extirpated from nature. Of those that were imperiled in 1989, most (89%) are the same or worse in conservation status; only 6% have improved in status, and 5% were delisted for various reasons. Habitat degradation and nonindigenous species are the main threats to at-risk fishes, many of which are restricted to small ranges. Documenting the diversity and status of rare fishes is a critical step in identifying and implementing appropriate actions necessary for their protection and management. Howard L. Jelks, Frank McCormick, Stephen J. Walsh, Joseph S. Nelson, Noel M. Burkhead, Steven P. Platania, Salvador Contreras-Balderas, Brady A. Porter, Edmundo Díaz-Pardo, Claude B. Renaud, Dean A. Hendrickson, Juan Jacobo Schmitter-Soto, John Lyons, Eric B. Taylor, and Nicholas E. Mandrak, Melvin L. Warren, Jr. Jelks, Walsh, and Burkhead are research McCormick is a biologist with the biologists with the U.S. -

ECOLOGY of NORTH AMERICAN FRESHWATER FISHES
ECOLOGY of NORTH AMERICAN FRESHWATER FISHES Tables STEPHEN T. ROSS University of California Press Berkeley Los Angeles London © 2013 by The Regents of the University of California ISBN 978-0-520-24945-5 uucp-ross-book-color.indbcp-ross-book-color.indb 1 44/5/13/5/13 88:34:34 AAMM uucp-ross-book-color.indbcp-ross-book-color.indb 2 44/5/13/5/13 88:34:34 AAMM TABLE 1.1 Families Composing 95% of North American Freshwater Fish Species Ranked by the Number of Native Species Number Cumulative Family of species percent Cyprinidae 297 28 Percidae 186 45 Catostomidae 71 51 Poeciliidae 69 58 Ictaluridae 46 62 Goodeidae 45 66 Atherinopsidae 39 70 Salmonidae 38 74 Cyprinodontidae 35 77 Fundulidae 34 80 Centrarchidae 31 83 Cottidae 30 86 Petromyzontidae 21 88 Cichlidae 16 89 Clupeidae 10 90 Eleotridae 10 91 Acipenseridae 8 92 Osmeridae 6 92 Elassomatidae 6 93 Gobiidae 6 93 Amblyopsidae 6 94 Pimelodidae 6 94 Gasterosteidae 5 95 source: Compiled primarily from Mayden (1992), Nelson et al. (2004), and Miller and Norris (2005). uucp-ross-book-color.indbcp-ross-book-color.indb 3 44/5/13/5/13 88:34:34 AAMM TABLE 3.1 Biogeographic Relationships of Species from a Sample of Fishes from the Ouachita River, Arkansas, at the Confl uence with the Little Missouri River (Ross, pers. observ.) Origin/ Pre- Pleistocene Taxa distribution Source Highland Stoneroller, Campostoma spadiceum 2 Mayden 1987a; Blum et al. 2008; Cashner et al. 2010 Blacktail Shiner, Cyprinella venusta 3 Mayden 1987a Steelcolor Shiner, Cyprinella whipplei 1 Mayden 1987a Redfi n Shiner, Lythrurus umbratilis 4 Mayden 1987a Bigeye Shiner, Notropis boops 1 Wiley and Mayden 1985; Mayden 1987a Bullhead Minnow, Pimephales vigilax 4 Mayden 1987a Mountain Madtom, Noturus eleutherus 2a Mayden 1985, 1987a Creole Darter, Etheostoma collettei 2a Mayden 1985 Orangebelly Darter, Etheostoma radiosum 2a Page 1983; Mayden 1985, 1987a Speckled Darter, Etheostoma stigmaeum 3 Page 1983; Simon 1997 Redspot Darter, Etheostoma artesiae 3 Mayden 1985; Piller et al. -

Spring 2021 Magazine Insert
KENTUCKY Spring 2021 • nature.org/kentucky Green River Lock and Dam #5 is slated for removal in fall 2021. © Mike Wilkinson Green River Lock and Dam #5 Biological assessment clears the way for dam removal Removing Green River Lock and Dam #5 mussel, to name just a few,” says Danna The assessment found that seven of at the Butler and Warren county line will Baxley, director of conservation for the the threatened and endangered species restore 200 miles of free-flowing con- Kentucky chapter. “Our consultants did will not be affected by the dam removal ditions to the river for the first time in a really deep assessment on where the process, while nine have the potential more than a century. Lock and Dam #5 species occurred and how the removal to be negatively impacted. The report is the linchpin in the largest river resto- process and changes in flow will affect outlines mitigation techniques needed to ration project in state history, and The those species.” protect those impacted species. Several Nature Conservancy, U.S. Army Corps The consultants evaluated conditions of the techniques focus on controlling of Engineers, and U.S. Fish and Wildlife both upstream and downstream of sediments and erosion. Service plan to commence removal efforts the dam and found that the 12.6 miles “The dam removal crew will be in the fall of 2021. Before moving forward upstream contained no habitat for any working during the lowest flows and with actual removal, however, TNC and of the mussel species. This is because building work paths that are not near its partners have to complete a myriad the water is pooled upstream of the dam, endangered species habitat,” Baxley says. -

Aquatic Fish Report
Aquatic Fish Report Acipenser fulvescens Lake St urgeon Class: Actinopterygii Order: Acipenseriformes Family: Acipenseridae Priority Score: 27 out of 100 Population Trend: Unknown Gobal Rank: G3G4 — Vulnerable (uncertain rank) State Rank: S2 — Imperiled in Arkansas Distribution Occurrence Records Ecoregions where the species occurs: Ozark Highlands Boston Mountains Ouachita Mountains Arkansas Valley South Central Plains Mississippi Alluvial Plain Mississippi Valley Loess Plains Acipenser fulvescens Lake Sturgeon 362 Aquatic Fish Report Ecobasins Mississippi River Alluvial Plain - Arkansas River Mississippi River Alluvial Plain - St. Francis River Mississippi River Alluvial Plain - White River Mississippi River Alluvial Plain (Lake Chicot) - Mississippi River Habitats Weight Natural Littoral: - Large Suitable Natural Pool: - Medium - Large Optimal Natural Shoal: - Medium - Large Obligate Problems Faced Threat: Biological alteration Source: Commercial harvest Threat: Biological alteration Source: Exotic species Threat: Biological alteration Source: Incidental take Threat: Habitat destruction Source: Channel alteration Threat: Hydrological alteration Source: Dam Data Gaps/Research Needs Continue to track incidental catches. Conservation Actions Importance Category Restore fish passage in dammed rivers. High Habitat Restoration/Improvement Restrict commercial harvest (Mississippi River High Population Management closed to harvest). Monitoring Strategies Monitor population distribution and abundance in large river faunal surveys in cooperation -
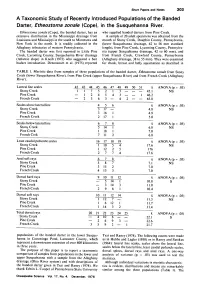
A Taxonomic Study of Recently Introduced Populations of The
Short Papers and Notes 303 A TaxonomicStudy of RecentlyIntroduced Populations of the Banded Darter,Etheostoma zonale (Cope), in the Susquehanna River. Etheostoma zonale (Cope), the banded darter, has an who supplied banded darters from Pine Creek. extensive distribution in the Mississippi drainage from A sample of 20 adult specimens was obtained from the Louisiana and Mississippi in the south to Minnesota and mouth of Stony Creek, Dauphin County, Pennsylvania, New York in the north. It is readily collected in the (lower Susquehanna drainage, 42 to 56 mm standard Allegheny tributaries of western Pennsylvania. length), from Pine Creek, Lycoming County, Pennsylva- The banded darter was first reported in Little Pine nia (upper Susquehanna drainage, 42 to 60 mm), and Creek, Lycoming County, Susquehanna River drainage from French Creek, Crawford County, Pennsylvania (Atlantic slope) in Kneib (1972) who suggested a bait (Allegheny drainage, 38 to 55 mm). They were examined bucket introduction. Denoncourt et al. (1975) reported for cheek, breast and belly squamation as described in TABLE 1. Meristic data from samples of three populations of the banded darter, Etheostoma zonale from Stony Creek (lower Susquehanna River), from Pine Creek (upper Susquehanna River) and from French Creek (Allegheny River). Lateral line scales 42 43 44 45 46 47 48 49 50 51 k ANOVA (p = .05) Stony Creek 1 1 7 5 2 1 3 - -- 45.1 NS Pine Creek - 1 2 4 5 4 3 - - 1 46.2 French Creek - 2 3 6 3 - 4 2 - - 45.8 Scales above lateral line 4 5 6 R ANOVA (p = .05) Stony Creek 3 -

DISTRIBUTION, ECOLOGY, and REPRODUCTIVE BIOLOGY of the ORANGEFIN MADTOM (NOTURUS GILBERTI) by Timothy Dale Simonson
DISTRIBUTION, ECOLOGY, AND REPRODUCTIVE BIOLOGY OF THE ORANGEFIN MADTOM (NOTURUS GILBERTI) by Timothy Dale Simonson Thesis submitted to the Faculty of the Virginia Polytechnic Institute and State University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE in Fisheries and Wildlife Sciences APPROVED: Richard J. Neves, Chair Dona:;[d J. Orth Johm J. Ney Louis A. Hel:frich April 1987 Blacksburg, Virginia DISTRIBUTION, ECOLOGY, AND REPRODUCTIVE BIOLOGY OF THE ORANGEFIN MADTOM (NOTURUS GILBERTI) by Timothy Dale Simonson Richard J. Neves, Chair Fisheries and Wildlife Sciences (ABSTRACT) Distribution of the orangefin madtom (Noturus gilberti) was determined from 347 sites sampled in Virginia and North Carolina. This species inhabited 264 stream kilometers, over twice the reported range, in the following systems: Craig Creek, Roanoke River, Dan River, Big Chestnut Creek, South Mayo River, Pigg River, and Smith River. The orangefin madtom was somewhat common; 33% (Dan River) to 70% (Craig Creek) of the sites sampled were occupied. Negative interspecific associates of orangefin madtoms included chubs, mountain redbelly dace, rosyside dace, crescent shiners, and crayfish; only Roanoke darters were considered positive associates. Sand and silt levels were significantly lower at sites with !L. gilberti, while per- centage of small cobble, local gradient, and depth were sig- nificantly higher. Discriminant function analysis identified large gravel, local gradient, silt, and occurrence of rosyside dace and crayfish, as significant predictors of the occurrence of the orangefin madtom. Seasonal samples from Craig Creek consisted of three age groups. The smallest individual captured was 33 mm total length (TL) and the largest was 111 mm TL. -

An Annotated Check-List of the Fishes of the Iowa-Cedar River Drainage Basin in Iowa," Proceedings of the Iowa Academy of Science: Vol
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by University of Northern Iowa Proceedings of the Iowa Academy of Science Volume 60 | Annual Issue Article 86 1953 An Annotated Check-List of the Fishes of the Iowa- Cedar River Drainage Basin in Iowa Robert E. Cleary Iowa Conservation Commission Copyright © Copyright 1953 by the Iowa Academy of Science, Inc. Follow this and additional works at: https://scholarworks.uni.edu/pias Recommended Citation Cleary, Robert E. (1953) "An Annotated Check-List of the Fishes of the Iowa-Cedar River Drainage Basin in Iowa," Proceedings of the Iowa Academy of Science: Vol. 60: No. 1 , Article 86. Available at: https://scholarworks.uni.edu/pias/vol60/iss1/86 This Research is brought to you for free and open access by UNI ScholarWorks. It has been accepted for inclusion in Proceedings of the Iowa Academy of Science by an authorized editor of UNI ScholarWorks. For more information, please contact [email protected]. Cleary: An Annotated Check-List of the Fishes of the Iowa-Cedar River Dra An Annotated Check-List of the Fishes of the Iowa-Cedar River Drainage Basin in Iowa By ROBERT E. CLEARY The Iowa-Cedar River drainage basin, which finds the larger of the two rivers, the Cedar, as a tributary to the smaller, is second in size of various inland river drainage basins in the state. The combined rivers drain 11,615 square miles or, roughly, one-fifth of the area of the state. The Iowa River, which has its source in Crystal Lake, in Han cock County, flows southeast for a distance of 329 miles and has a 685-foot fall from source to mouth. -

Kansas Stream Fishes
A POCKET GUIDE TO Kansas Stream Fishes ■ ■ ■ ■ ■ ■ ■ ■ ■ ■ By Jessica Mounts Illustrations © Joseph Tomelleri Sponsored by Chickadee Checkoff, Westar Energy Green Team, Kansas Department of Wildlife, Parks and Tourism, Kansas Alliance for Wetlands & Streams, and Kansas Chapter of the American Fisheries Society Published by the Friends of the Great Plains Nature Center Table of Contents • Introduction • 2 • Fish Anatomy • 3 • Species Accounts: Sturgeons (Family Acipenseridae) • 4 ■ Shovelnose Sturgeon • 5 ■ Pallid Sturgeon • 6 Minnows (Family Cyprinidae) • 7 ■ Southern Redbelly Dace • 8 ■ Western Blacknose Dace • 9 ©Ryan Waters ■ Bluntface Shiner • 10 ■ Red Shiner • 10 ■ Spotfin Shiner • 11 ■ Central Stoneroller • 12 ■ Creek Chub • 12 ■ Peppered Chub / Shoal Chub • 13 Plains Minnow ■ Silver Chub • 14 ■ Hornyhead Chub / Redspot Chub • 15 ■ Gravel Chub • 16 ■ Brassy Minnow • 17 ■ Plains Minnow / Western Silvery Minnow • 18 ■ Cardinal Shiner • 19 ■ Common Shiner • 20 ■ Bigmouth Shiner • 21 ■ • 21 Redfin Shiner Cover Photo: Photo by Ryan ■ Carmine Shiner • 22 Waters. KDWPT Stream ■ Golden Shiner • 22 Survey and Assessment ■ Program collected these Topeka Shiner • 23 male Orangespotted Sunfish ■ Bluntnose Minnow • 24 from Buckner Creek in Hodgeman County, Kansas. ■ Bigeye Shiner • 25 The fish were catalogued ■ Emerald Shiner • 26 and returned to the stream ■ Sand Shiner • 26 after the photograph. ■ Bullhead Minnow • 27 ■ Fathead Minnow • 27 ■ Slim Minnow • 28 ■ Suckermouth Minnow • 28 Suckers (Family Catostomidae) • 29 ■ River Carpsucker • -

A Distributional Atlas of Upper Mississippi River Fishes
LIBRARY OF THE UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN Y\o. GG - ^9 SURVEY ; (26814—2M—2-71 A DISTRIBUTIONAL ATLAS OF UPPER MISSISSIPPI RIVER FISHES PHILIP W. SMITH \LVIN C. LOPINOT WILLIAM L. PFLIEGER STATE OF ILLINOIS ISIOLOGICAL NOTES NO. 73 DEPARTMENT OF REGISTRATION AND EDUCATION ILINOIS NATURAL HISTORY SURVEY NATURAL HISTORY SURVEY DIVISION iJRBANA, ILLINOIS • MAY, 1971 Minn. numbers refer to navigation pools and their downstream locks Fig. 1 .—The upper Mississippi River and its major tributaries. The referred to as B-26. and dams. Note the absence of a pool 23 and of locks and dams below dam 26 in the unimpounded river, A DISTRIBUTIONAL ATLAS OF UPPER MISSISSIPPI RIVER FISHES Philip W. Smith, Alvin C. Lopinot, and William L. Pflieger GROWING CONCERN OVER POLLUTION and in the Museum of Zoology at the University of Michigan, habitat alteration in the Mississippi Ri\er prompts us old reports of federal and state fish commissions and the to publish information we have assembled on the present U.S. Bureau of Fisheries, and scattered publications citing distribution of fishes in that river. Since 1962 a concerted records from the Mississippi River. For an excellent efTort, under the aegis of the Upper Mississippi River summary of early investigations, see Carlander (1954). Conservation Committee (hereafter UMRCC), has re- With the establishment of the UMRCC, its Fish sulted in a vast quantity of distributional data, and its Technical Section inaugurated field operations early in publication can provide a criterion that will aid in the 1944. These investigations continued through 1946 and detection and measurement of future changes in the en- involved personnel and financial support from the con- vironmental quality of the river.