Botany Basics ❂1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Taxonomic Status of Gladiolus Illyricus (Iridaceae) in Britain
The Taxonomic Status of Gladiolus illyricus (Iridaceae) in Britain Aeron Buchanan Supervisor: Fred Rumsey, Natural History Museum, London A thesis submitted in partial fulfilment of the requirements for the degree of Master of Science of Imperial College, London Abstract First noticed officially in Britain in 1855, Gladiolus illyricus (Koch) presents an interesting taxonomic and biogeographical challenge: whether or not this isolated northern population should be recognized as a separate sub-species. Fundamental conservation issues rest on the outcome. Here, the investigation into the relationship of the G. illyricus plants of the New Forest, Hampshire, to Gladiolus species across Europe, northern Africa and the middle east is initiated. Two chloroplast regions, one in trnL–trnF and the other across psbA–trnH have been sequenced for 42 speci- mens of G. illyricus, G. communis, G. italicus, G. atroviolaceus, G. triphyllos and G. anatolicus. Phylogenetic and biogeographical treatments support the notion of an east–west genetic gradation along the Mediterranean. Iberia particularly appears as a zone of high hybridization potential and the source of the New Forest population. Alignment with sequences obtained from GenBank give strong support to the classic taxonomy of Gladiolus being monophyletic in its sub-family, Ixioideae. Comments on these chloroplast regions for barcoding are also given. In conclusion, the genetic localization of Britain’s G. illyricus population as an extremity haplotype suggests that it could well deserve sub-species status. Contents 1 Introduction 2 2 Background 4 3 Materials and Methods 8 4 Results and Discussion 15 5 Conclusions 26 Appendices 28 References 56 1. Introduction G. illyricus in Britain Figure 1: G. -

Invasive Weeds of the Appalachian Region
$10 $10 PB1785 PB1785 Invasive Weeds Invasive Weeds of the of the Appalachian Appalachian Region Region i TABLE OF CONTENTS Acknowledgments……………………………………...i How to use this guide…………………………………ii IPM decision aid………………………………………..1 Invasive weeds Grasses …………………………………………..5 Broadleaves…………………………………….18 Vines………………………………………………35 Shrubs/trees……………………………………48 Parasitic plants………………………………..70 Herbicide chart………………………………………….72 Bibliography……………………………………………..73 Index………………………………………………………..76 AUTHORS Rebecca M. Koepke-Hill, Extension Assistant, The University of Tennessee Gregory R. Armel, Assistant Professor, Extension Specialist for Invasive Weeds, The University of Tennessee Robert J. Richardson, Assistant Professor and Extension Weed Specialist, North Caro- lina State University G. Neil Rhodes, Jr., Professor and Extension Weed Specialist, The University of Ten- nessee ACKNOWLEDGEMENTS The authors would like to thank all the individuals and organizations who have contributed their time, advice, financial support, and photos to the crea- tion of this guide. We would like to specifically thank the USDA, CSREES, and The Southern Region IPM Center for their extensive support of this pro- ject. COVER PHOTO CREDITS ii 1. Wavyleaf basketgrass - Geoffery Mason 2. Bamboo - Shawn Askew 3. Giant hogweed - Antonio DiTommaso 4. Japanese barberry - Leslie Merhoff 5. Mimosa - Becky Koepke-Hill 6. Periwinkle - Dan Tenaglia 7. Porcelainberry - Randy Prostak 8. Cogongrass - James Miller 9. Kudzu - Shawn Askew Photo credit note: Numbers in parenthesis following photo captions refer to the num- bered photographer list on the back cover. HOW TO USE THIS GUIDE Tabs: Blank tabs can be found at the top of each page. These can be custom- ized with pen or marker to best suit your method of organization. Examples: Infestation present On bordering land No concern Uncontrolled Treatment initiated Controlled Large infestation Medium infestation Small infestation Control Methods: Each mechanical control method is represented by an icon. -

Physicochemical Properties of Selected Root and Tuber Starches
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1999 Physicochemical properties of selected root and tuber starches and characterization of extruded, chemically modified corn starches Andrew Edward McPherson Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Agricultural Science Commons, Agriculture Commons, Agronomy and Crop Sciences Commons, Food Science Commons, and the Plant Biology Commons Recommended Citation McPherson, Andrew Edward, "Physicochemical properties of selected root and tuber starches and characterization of extruded, chemically modified corn starches " (1999). Retrospective Theses and Dissertations. 12592. https://lib.dr.iastate.edu/rtd/12592 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI fihns the text directly from the ori^nal or copy submitted. Thus, some thesis and dissertation copies are in typewriter &ce, v^e others may be from any type of computo^ printer. The quality of this reproduction is dependeut upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely a£fect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. -

Evaluating Ethylene Sensitivity Using Mature Plant Screens and the Seedling Hypocotyl Response
Evaluating Ethylene Sensitivity Using Mature Plant Screens and the Seedling Hypocotyl Response THESIS Presented in Partial Fulfillment of the Requirements for the Degree Master of Science in the Graduate School of The Ohio State University By Nichole Frances Edelman, B. A. Graduate Program in Horticulture and Crop Science The Ohio State University 2013 Master's Examination Committee: Dr. Michelle L. Jones, Advisor Dr. Pablo Jourdan Dr. Claudio Pasian Copyrighted by Nichole Frances Edelman 2013 Abstract Ethylene (C2H4) is a gaseous hormone produced by plants in response to environmental stress and during growth and development. Sensitive seedlings exposed to ethylene gas will exhibit an exaggerated apical hook, thickened hypocotyl, and reduced hypocotyl elongation. Collectively these symptoms are known as the triple response, and have been used as a screen to identify ethylene mutants. Exposure to ethylene gas at the mature plant stage can induce flower, bud, or leaf abscission, flower senescence, leaf chlorosis (yellowing), or leaf epinasty (downward curvature). Sensitivity can vary between species and by developmental stage or tissue (flower vs leaf). Ethylene can damage plants during production, shipping, and retailing. The objectives of this research were 1) to determine if the seedling hypocotyl elongation screen could be used to predict the sensitivity of mature plants at the marketable stage; 2) to identify ethylene sensitivity differences (levels of sensitivity and symptoms) between accessions within Solanaceae; and 3) to identify ethylene sensitivity differences at different developmental stages (seedling, juvenile, and mature plants). Seedlings were germinated on filter paper saturated with 1- aminocyclopropane-1-carboxylic acid (ACC) and sensitivity was determined based on the hypocotyl lengths relative to the control (0 µM ACC). -

Growing the Gladiolus
Volume 3 Article 1 Bulletin P85 Growing the Gladiolus 7-1-1947 Growing the Gladiolus L. C. Grove Iowa State College Follow this and additional works at: http://lib.dr.iastate.edu/bulletinp Part of the Agriculture Commons, and the Plant Sciences Commons Recommended Citation Grove, L. C. (1947) "Growing the Gladiolus," Bulletin P: Vol. 3 : Bulletin P85 , Article 1. Available at: http://lib.dr.iastate.edu/bulletinp/vol3/iss85/1 This Article is brought to you for free and open access by the Iowa Agricultural and Home Economics Experiment Station Publications at Iowa State University Digital Repository. It has been accepted for inclusion in Bulletin P by an authorized editor of Iowa State University Digital Repository. For more information, please contact [email protected]. Grove: Growing the Gladiolus g N SIO N SER V IC E, Cooperating Published by Iowa State University Digital Repository, 1947 AMES, IOWA 1 Bulletin P, Vol. 3, No. 85 [1947], Art. 1 CONTENTS Page Growth development ....................................................................,807 Propagation ................................ .. ........................................... 809 Methods ..................................................... 809 Bulbs . , .......................................................... 810 Sizes ......................................•.....................................................810 Bulb shape ............................................................. 810 Varieties .............................. 811 Flower uses ................. ...................................................................811 -

Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy
plants Article Ongoing Evolution in the Genus Crocus: Diversity of Flowering Strategies on the Way to Hysteranthy Teresa Pastor-Férriz 1, Marcelino De-los-Mozos-Pascual 1, Begoña Renau-Morata 2, Sergio G. Nebauer 2 , Enrique Sanchis 2, Matteo Busconi 3 , José-Antonio Fernández 4, Rina Kamenetsky 5 and Rosa V. Molina 2,* 1 Departamento de Gestión y Conservación de Recursos Fitogenéticos, Centro de Investigación Agroforestal de Albadaledejito, 16194 Cuenca, Spain; [email protected] (T.P.-F.); [email protected] (M.D.-l.-M.-P.) 2 Departamento de Producción Vegetal, Universitat Politècnica de València, 46022 Valencia, Spain; [email protected] (B.R.-M.); [email protected] (S.G.N.); [email protected] (E.S.) 3 Department of Sustainable Crop Production, Università Cattolica del Sacro Cuore, 29122 Piacenza, Italy; [email protected] 4 IDR-Biotechnology and Natural Resources, Universidad de Castilla-La Mancha, 02071 Albacete, Spain; [email protected] 5 Department of Ornamental Horticulture and Biotechnology, The Volcani Center, ARO, Rishon LeZion 7505101, Israel; [email protected] * Correspondence: [email protected] Abstract: Species of the genus Crocus are found over a wide range of climatic areas. In natural habitats, these geophytes diverge in the flowering strategies. This variability was assessed by analyzing the flowering traits of the Spanish collection of wild crocuses, preserved in the Bank of Plant Germplasm Citation: Pastor-Férriz, T.; of Cuenca. Plants of the seven Spanish species were analyzed both in their natural environments De-los-Mozos-Pascual, M.; (58 native populations) and in common garden experiments (112 accessions). -

Effects of Nitrogen Dioxide on Biochemical Responses in 41 Garden Plants
plants Article Effects of Nitrogen Dioxide on Biochemical Responses in 41 Garden Plants Qianqian Sheng 1 and Zunling Zhu 1,2,* 1 College of Landscape Architecture, Nanjing Forestry University, Nanjing 210037, China; [email protected] 2 College of Art & Design, Nanjing Forestry University, Nanjing 210037, China * Correspondence: [email protected]; Tel.: +86-25-6822-4603 Received: 11 December 2018; Accepted: 12 February 2019; Published: 16 February 2019 Abstract: Nitrogen dioxide (NO2) at a high concentration is among the most common and harmful air pollutants. The present study aimed to explore the physiological responses of plants exposed to NO2. A total of 41 plants were classified into 13 functional groups according to the Angiosperm Phylogeny Group classification system. The plants were exposed to 6 µL/L NO2 in an open-top glass chamber. The physiological parameters (chlorophyll (Chl) content, peroxidase (POD) activity, and soluble protein and malondialdehyde (MDA) concentrations) and leaf mineral ion contents (nitrogen (N+), phosphorus (P+), potassium (K+), calcium (Ca2+), magnesium (Mg2+), manganese 2+ 2+ (Mn ), and zinc (Zn )) of 41 garden plants were measured. After NO2 exposure, the plants were subsequently transferred to a natural environment for a 30-d recovery to determine whether they could recover naturally and resume normal growth. The results showed that NO2 polluted the plants and that NO2 exposure affected leaf Chl contents in most functional groups. Increases in both POD activity and soluble protein and MDA concentrations as well as changes in mineral ion concentrations could act as signals for inducing defense responses. Furthermore, antioxidant status played an important role in plant protection against NO2-induced oxidative damage. -

The Extent and Genetic Basis of Phenotypic Divergence in Life History Traits in Mimulus Guttatus
Molecular Ecology (2014) doi: 10.1111/mec.13004 The extent and genetic basis of phenotypic divergence in life history traits in Mimulus guttatus JANNICE FRIEDMAN,* ALEX D. TWYFORD,*† JOHN H. WILLIS‡ and BENJAMIN K. BLACKMAN§ *Department of Biology, Syracuse University, 110 College Place, Syracuse, NY 13244, USA, †Institute of Evolutionary Biology, University of Edinburgh, Mayfield Rd., Edinburgh EH9 3JT, UK, ‡Department of Biology, Duke University, Box 90338, Durham, NC 27708, USA, §Department of Biology, University of Virginia, Box 400328, Charlottesville, VA 22904, USA Abstract Differential natural selection acting on populations in contrasting environments often results in adaptive divergence in multivariate phenotypes. Multivariate trait divergence across populations could be caused by selection on pleiotropic alleles or through many independent loci with trait-specific effects. Here, we assess patterns of association between a suite of traits contributing to life history divergence in the common monkey flower, Mimulus guttatus, and examine the genetic architecture underlying these corre- lations. A common garden survey of 74 populations representing annual and perennial strategies from across the native range revealed strong correlations between vegetative and reproductive traits. To determine whether these multitrait patterns arise from pleiotropic or independent loci, we mapped QTLs using an approach combining high- throughput sequencing with bulk segregant analysis on a cross between populations with divergent life histories. We find extensive pleiotropy for QTLs related to flower- ing time and stolon production, a key feature of the perennial strategy. Candidate genes related to axillary meristem development colocalize with the QTLs in a manner consistent with either pleiotropic or independent QTL effects. Further, these results are analogous to previous work showing pleiotropy-mediated genetic correlations within a single population of M. -

The Vascular System of Monocotyledonous Stems Author(S): Martin H
The Vascular System of Monocotyledonous Stems Author(s): Martin H. Zimmermann and P. B. Tomlinson Source: Botanical Gazette, Vol. 133, No. 2 (Jun., 1972), pp. 141-155 Published by: The University of Chicago Press Stable URL: http://www.jstor.org/stable/2473813 . Accessed: 30/08/2011 15:50 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to Botanical Gazette. http://www.jstor.org 1972] McCONNELL& STRUCKMEYER ALAR AND BORON-DEFICIENTTAGETES 141 tomato, turnip and cotton to variations in boron nutri- Further investigationson the relation of photoperiodto tion. II. Anatomical responses. BOT.GAZ. 118:53-71. the boron requirementsof plants. BOT.GAZ. 109:237-249. REED, D. J., T. C. MOORE, and J. D. ANDERSON. 1965. Plant WATANABE,R., W. CHORNEY,J. SKOK,and S. H. WENDER growth retardant B-995: a possible mode of action. 1964. Effect of boron deficiency on polyphenol produc- Science 148: 1469-1471. tion in the sunflower.Phytochemistry 3:391-393. SKOK, J. 1957. Relationships of boron nutrition to radio- ZEEVAART,J. A. D. 1966. Inhibition of stem growth and sensitivity of sunflower plants. -
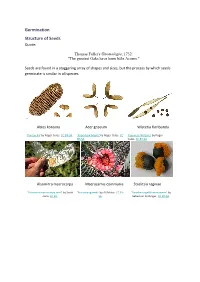
Germination Structure of Seeds Quote
Germination Structure of Seeds Quote: Thomas Fuller's Gnomologia, 1732: "The greatest Oaks have been little Acorns." Seeds are found in a staggering array of shapes and sizes, but the process by which seeds germinate is similar in all species. Abies koreana Acer griseum Wisteria floribunda 'Korean Fir' by Roger Culos. CC BY-SA. 'Paperbark Maple' by Roger Culos. CC 'Japanese Wisteria' by Roger BY-SA. Culos. CC BY-SA. Alsomitra macrocarpa Macrozamia communis Strelitzia reginae 'Alsomitra macrocarpa seed' by Scott 'BurrawangSeeds' by AYArktos. CC BY- 'Paradiesvogelblumensamen' by Zona. CC BY. SA. Sebastian Stabinger. CC BY-SA. Taraxicum officinale Stephanotis floribunda Phleum pratense 'Achane of Taraxacum sect. 'Stephanotis seed' by L. Marie"/Lenore 'Timoteegras vruchten Phleum Ruderalia' by Didier Edman, Sunnyvale, CA. CC BY. pratense' by Rasbak. CC BY-SA. Descouens. CC BY-SA. Dicotyledon seeds testa epicotyl plumule hypocotyl cotyledon radicle 'Aesculus hippocastanum seed section' by Boronian. CC BY. plumule epicotyl hypocotyl testa hilum radicle cotyledon micropyle endosperm Monocotyledon seeds endosperm epicotyl testa hypocotyl cotyledon radicle Parts of a seed Testa The seed coat. A protective layer which is tough and hard and it protects the seed from attack by insects, fungi and bacteria. Cotyledon Dicotyledons have 2 cotyledons Monocotyledons have 1 cotyledon A cotyledon is an embryonic leaf. It is the first leaf to appear when a seedling grows. They often contain reserves of food which the developing seedling can use to grow. Epicotyl The section of stem between the cotyledon(s) and the plumule. In a seedling it is the section of stem between the cotyledons and the first true leaves. -

Chapter 1 Definitions and Classifications for Fruit and Vegetables
Chapter 1 Definitions and classifications for fruit and vegetables In the broadest sense, the botani- Botanical and culinary cal term vegetable refers to any plant, definitions edible or not, including trees, bushes, vines and vascular plants, and Botanical definitions distinguishes plant material from ani- Broadly, the botanical term fruit refers mal material and from inorganic to the mature ovary of a plant, matter. There are two slightly different including its seeds, covering and botanical definitions for the term any closely connected tissue, without vegetable as it relates to food. any consideration of whether these According to one, a vegetable is a are edible. As related to food, the plant cultivated for its edible part(s); IT botanical term fruit refers to the edible M according to the other, a vegetable is part of a plant that consists of the the edible part(s) of a plant, such as seeds and surrounding tissues. This the stems and stalk (celery), root includes fleshy fruits (such as blue- (carrot), tuber (potato), bulb (onion), berries, cantaloupe, poach, pumpkin, leaves (spinach, lettuce), flower (globe tomato) and dry fruits, where the artichoke), fruit (apple, cucumber, ripened ovary wall becomes papery, pumpkin, strawberries, tomato) or leathery, or woody as with cereal seeds (beans, peas). The latter grains, pulses (mature beans and definition includes fruits as a subset of peas) and nuts. vegetables. Definition of fruit and vegetables applicable in epidemiological studies, Fruit and vegetables Edible plant foods excluding -

AXR1 Acts After Lateral Bud Formation to Inhibit Lateral Bud Growth in Arabidopsis
This is a repository copy of AXR1 acts after lateral bud formation to inhibit lateral bud growth in Arabidopsis. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/262/ Article: Stirnberg, P., Leyser, O. and Chatfield, S.P. (1999) AXR1 acts after lateral bud formation to inhibit lateral bud growth in Arabidopsis. Plant Physiology. pp. 839-847. ISSN 0032-0889 Reuse Items deposited in White Rose Research Online are protected by copyright, with all rights reserved unless indicated otherwise. They may be downloaded and/or printed for private study, or other acts as permitted by national copyright laws. The publisher or other rights holders may allow further reproduction and re-use of the full text version. This is indicated by the licence information on the White Rose Research Online record for the item. Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Plant Physiology, November 1999, Vol. 121, pp. 839–847, www.plantphysiol.org © 1999 American Society of Plant Physiologists AXR1 Acts after Lateral Bud Formation to Inhibit Lateral Bud Growth in Arabidopsis1 Petra Stirnberg, Steven P. Chatfield, and H.M. Ottoline Leyser* Department of Biology, University of York, P.O. Box 373, York YO10 5YW, United Kingdom Several mutants with altered auxin sensitivity have been The AXR1 gene of Arabidopsis is required for many auxin re- produced in Arabidopsis.