Wurmbea Dioica
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Tracing History
Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 911 Tracing History Phylogenetic, Taxonomic, and Biogeographic Research in the Colchicum Family BY ANNIKA VINNERSTEN ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2003 Dissertation presented at Uppsala University to be publicly examined in Lindahlsalen, EBC, Uppsala, Friday, December 12, 2003 at 10:00 for the degree of Doctor of Philosophy. The examination will be conducted in English. Abstract Vinnersten, A. 2003. Tracing History. Phylogenetic, Taxonomic and Biogeographic Research in the Colchicum Family. Acta Universitatis Upsaliensis. Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 911. 33 pp. Uppsala. ISBN 91-554-5814-9 This thesis concerns the history and the intrafamilial delimitations of the plant family Colchicaceae. A phylogeny of 73 taxa representing all genera of Colchicaceae, except the monotypic Kuntheria, is presented. The molecular analysis based on three plastid regions—the rps16 intron, the atpB- rbcL intergenic spacer, and the trnL-F region—reveal the intrafamilial classification to be in need of revision. The two tribes Iphigenieae and Uvularieae are demonstrated to be paraphyletic. The well-known genus Colchicum is shown to be nested within Androcymbium, Onixotis constitutes a grade between Neodregea and Wurmbea, and Gloriosa is intermixed with species of Littonia. Two new tribes are described, Burchardieae and Tripladenieae, and the two tribes Colchiceae and Uvularieae are emended, leaving four tribes in the family. At generic level new combinations are made in Wurmbea and Gloriosa in order to render them monophyletic. The genus Androcymbium is paraphyletic in relation to Colchicum and the latter genus is therefore expanded. -

Ecological Differentiation of Combined and Separate Sexes of Wurmbea Dioica (Colchicaceae) in Sympatry
Ecology, 82(9), 2001, pp. 2601±2616 q 2001 by the Ecological Society of America ECOLOGICAL DIFFERENTIATION OF COMBINED AND SEPARATE SEXES OF WURMBEA DIOICA (COLCHICACEAE) IN SYMPATRY ANDREA L. CASE1 AND SPENCER C. H. BARRETT Department of Botany, University of Toronto, 25 Willcocks Street, Toronto, Ontario, Canada M5S 3B2 Abstract. The evolution and maintenance of combined vs. separate sexes in ¯owering plants is in¯uenced by both ecological and genetic factors; variation in resources, partic- ularly moisture availability, is thought to play a role in selection for gender dimorphism in some groups. We investigated the density, distribution, biomass allocation, and physi- ology of sympatric monomorphic (cosexual) and dimorphic (female and male) populations of Wurmbea dioica in relation to soil moisture on the Darling Escarpment in southwestern Australia. Populations with monomorphic vs. dimorphic sexual systems segregated into wet vs. dry microsites, respectively, and biomass allocation patterns and physiological traits re¯ected differences in water availability, despite similarities in total ramet biomass between the sexual systems. Unisexuals ¯owered earlier at lower density, and they allocated sig- ni®cantly more biomass below ground to roots and corms than did cosexuals, which al- located more biomass above ground to leaves, stems, and ¯owers. Females, males, and cosexuals produced similar numbers of ¯owers per ramet, but unisexuals produced more ramets than cosexuals, increasing the total number of ¯owers per genet. Contrary to ex- pectation, cosexuals had signi®cantly higher (more positive) leaf carbon isotope ratios and lower leaf nitrogen content than unisexuals, suggesting that cosexuals are more water-use ef®cient and have lower rates of photosynthesis per unit leaf mass despite their occurrence in wetter microsites. -

The Species of Wurmbea
J. Adelaide Bot. Gard. 16: 33-53 (1995) THE SPECIES OFWURMBEA(LILIACEAE) IN SOUTH AUSTRALIA Robert J. Bates Cl- State Herbarium, Botanic Gardens, North Terrace, Adelaide, South Australia 5000 Abstract Nine species of Wunnbea Thunb. are recognised in South Australia. W. biglandulosa (R. Br.)Macfarlane, W. deserticola Macfarlane and W. sinora Macfarlane are recorded for the first time; Wurmbea biglandulosa ssp. flindersica, W. centralis ssp. australis, W. decumbens, W. dioica ssp. citrina, W. dioica ssp. lacunaria, W. latifolia ssp. vanessae and W. stellata are described. A key, together with notes on each species is provided. Macfarlane (1980) revised the genus for Australia. He placed Anguillaria R. Br. under Wurmbea and recognised W. dioica (R. Br.)F. Muell., W. centralis Macfarlane, W. latifolia Macfarlane and W. uniflora (R. Br.)Macfarlane as occurring in South Australia. Before this only one species, W. dioica (as Anguillaria dioica) was listed for South Australia (J.M. Black 1922, 1943). Macfarlane stated that he had seen no live material of South Australian species. The present author has made extensive field studies of taxa discussed in this paper, has cultivated most and studied herbarium material. Several trips have been made to other states to allow further comparisons to be made. For information on the nomenclatural history, general morphology, biology and ecology of Wurmbea see Macfarlane 1980. Key to the South Australian species of Wurmbea 1 Lower leaves paired (almost opposite), basal, of same shape and size 2 1: Lower leaves well separated, often of different shape and size 4 2 Leaves with serrate margins, flowers unisexual, nectaries 1 per tepal, a single band of colour... -
Native Vegetation of the Goulburn Broken Riverine Plains
Native Vegetation of the Goulburn Broken Riverine Plains Native Vegetation of the Goulburn Broken Riverine Plains This project is delivered and funded primarily through the partnerships between the Goulburn Broken Catchment Management Authority (GBCMA), Department of Primary Industries (DPI), Goulburn Murray Landcare Network (GMLN), Greater Shepparton City Council, Shire of Campaspe and Moira Shire. Published by: Goulburn Broken Catchment Management Authority 168 Welsford St, Shepparton, Victoria, Australia August 2012 ISBN: 978-1-920742-25-6 Acknowledgments The Goulburn Broken CMA and the GMLN gratefully acknowledge the staff of the Sustainable Irrigated Landscapes - Goulburn Broken, Environmental Management Team, particularly Fiona Copley who compiled the first edition “Native Vegetation in the Shepparton Irrigation Region” based on research of literature (References page 95) and communication with recognised flora scientists. Special acknowledgement goes to the GMLN in partnership with the Shepparton Irrigation Region Implementation Committee for enabling the printing of the first edition. The second edition, renamed “Native Vegetation of the Goulburn Broken Riverine Plains” was updated by Wendy D’Amore, GMLN with additions and subtractions made to the plant list and the booklet published in a new format. Special thanks to Sharon Terry, Rolf Weber, Joel Pyke and Gary Deayton for their expert knowledge of the plants and their distribution in the Riverine Plains. Many thanks also to members of the GMLN, Goulburn Broken CMA, DPI and Goulburn Valley Printing Services for their advice and assistance. Photo credits In this edition many plant profiles had their photographs updated or added to and additional species were added. The following photographers are gratefully acknowledged: Sharon Terry, Phil Hunter, Judy Ormond, Andrew Pearson, Keith Ward, Janet Hagen, Gary Deayton, Danielle Beischer, Bruce Wehner and Wendy D’Amore. -

A Review of South Australian Wurmbea (Colchicaceae~Liliaceae): Keys, New Taxa and Combinations, and Notes Robert J
© 2007 Board of the Botanic Gardens & State Herbarium, Government of South Australia J. Adelaide Bot. Gard. 21 (2007) 75–81 © 2007 Department for Environment & Heritage, Government of South Australia A review of South Australian Wurmbea (Colchicaceae~Liliaceae): keys, new taxa and combinations, and notes Robert J. Bates State Herbarium of South Australia, Hackney Road, Adelaide, South Australia 5000 Abstract Wurmbea nilpinna R.J.Bates, a rare local endemic in the Davenport Range on Nilpinna Station in the Lake Eyre region of South Australia, and W. dioica ssp. brevifolia R.J.Bates, widespread in the wheat belt of south-eastern Australia, are described as new and illustrated. W. citrina, based on W. dioica ssp. citrina R.J.Bates, and W. australism based on W. centralis ssp. australis R.J.Bates, are raised to species level. A key is provided for all the South Australian species together with additional distribution records and other notes. Introduction 3: Lower leaves narrow, linear, decumbent; flower single, less than 7 mm across; capsule elongate on a decumbent Prior to 1980 only one species of Wurmbea was stem . .W. decumbens recognised for South Australia (e.g., Black 1943). Lower leaves broadly linear-lanceolate, not decumbent; This was W. dioica (R.Br.)F.Muell., known then as flowers usually several, greater than 7 mm across; capsule Anguillaria dioica R.Br. ovoid on an erect stem . W. australis Macfarlane (1980) reduced the Australian genus 4: Nectary one per tepal, either central or as a single band of Anguillaria R. Br., the ‘early nancies’, to synonymy colour . 5 under Wurmbea and recognised four species from South Nectaries two per tepal . -

Colchicaceae) in Western Australia
Int. J. Plant Sci. 165(2):289–301. 2004. Ó 2004 by The University of Chicago. All rights reserved. 1058-5893/2004/16502-0007$15.00 FLORAL BIOLOGY OF GENDER MONOMORPHISM AND DIMORPHISM IN WURMBEA DIOICA (COLCHICACEAE) IN WESTERN AUSTRALIA Andrea Case1 and Spencer C. H. Barrett Department of Botany, University of Toronto, 25 Willcocks Street, Toronto, Ontario M5S 3B2, Canada We compared the floral biology of female, male, and cosexual plants in sympatric populations of Wurmbea dioica (Colchicaceae) from Western Australia to assess evidence that changes in pollination biology accompanied a transition between monomorphic to dimorphic sexual systems in this species. In W. dioica, sex- specific differences in floral design and display, the quantity and quality of rewards, and reproductive phenology represent alternative strategies for pollinator attraction and the avoidance of self-pollination. Unisexual plants had smaller flowers containing less pollen than flowers of cosexuals; males and cosexuals produced nectar with higher sugar content than did females. These patterns were associated with differences in the composition and visitation rates of pollinators. Nectar-foraging flies visited all three sexes, but pollen- collecting bees visited only cosexuals. Fly-pollinated females achieved greater seed fertility relative to their bee- pollinated cosexual relatives, and males and cosexuals received longer and more frequent visits than did females. Although there were no differences in flower number among sexes, females maintained larger displays throughout flowering. In contrast, males and cosexuals staggered both floral anthesis and anther dehiscence as mechanisms for promoting enhanced pollen dispersal. We propose that sex-based differences in floral design and display in W. -

Wurmbea Fluviatilis (Colchicaceae), a New Riverine Species 25
T.D.Nuytsia Macfarlane 21(1): 25–30 & A.L. (2011) Case, Wurmbea fluviatilis (Colchicaceae), a new riverine species 25 Wurmbea fluviatilis (Colchicaceae), a new riverine species from the Gascoyne region of Western Australia Terry D. Macfarlane1 and Andrea L. Case2 1Western Australian Herbarium. Postal address: Science Division, Department of Environment and Conservation, Locked Bag 2, Manjimup, Western Australia 6258 2Department of Biological Sciences, Kent State University, Kent OH 44242, USA Abstract Macfarlane, T.D. & Case, A.L. Wurmbea fluviatilis (Colchicaceae), a new riverine species from the Gascoyne region of Western Australia. Nuytsia 21(1): 25–30 (2011). A new species of Wurmbea, W. fluviatilisT.Macfarlane & A.Case, is described and illustrated with photographs and a distribution map. The new species is known from only three populations from the Gascoyne River catchment in the region of Mount Augustus, growing on river banks and beside riverside pools. It is a relatively tall, attractive species with bi-coloured flowers. Introduction A number of new species of Wurmbea Thunb. (Colchicaceae) have been discovered in Western Australia in recent years, most of them in the more arid regions outside of the better-collected south -west of the State. Following on from the description of W. inflata (Macfarlane & Case 2007), this is the second species to be described recently from the Gascoyne region. Wurmbea has been extensively studied taxonomically in Australia (Macfarlane 1980, 1987; Bates 1995, 2007) so the recent surge in discoveries is surprising, but is a consequence of targeted field work in under-collected areas at appropriate times. The new species described here was first collected during unrelated fieldwork by R.J. -
Lorenzo Peruzzi Male Flowers in Liliaceae Are More Frequent Than
Lorenzo Peruzzi Male flowers in Liliaceae are more frequent than previously thought Abstract Peruzzi, L.: Male flowers in Liliaceae are more frequent than previously thought. — Bocconea 24: 301-304. 2012. — ISSN 1120-4060. In the last twenty years a growing number of studies emphasized the occurrence of female-ster- ile reproductive systems within the monocot order Liliales. The occurrence of male flowers is here documented for the first time in Fritillaria involucrata, F. messanensis, F. montana, F. per- sica, Lilium bulbiferum subsp. croceum and Tulipa sylvestris. Increasingly frequent observa- tions of female-sterile systems within the order, and particularly in Liliaceae, suggest they could have an evolutionary significance. Introduction The adoption of sub-dioecious and sub-monoecious sexual models is rare among angiosperms (Charlesworth 2002). In particular, the female-sterile reproductive systems – andromonoecy and androdioecy – are considered the rarest strategies, being known for about 4,000 angiosperm species (Vallejo-Marín & Rausher 2007), which are approximate- ly 1% of the total number. Despite the general rarity of female-sterile reproductive systems, even including particular cases such as gender diphasy, in the last twenty years a growing number of studies are emphasizing the occurrence of these strategies within the monocot order Liliales, where about 20% of the ca. 1,600 species are known to be dioecious (i.e. the whole family Smilacaceae, Kong & al. 2007; Chamaelirium luteum – Melianthaceae, Meagher & Thompson 1987; Smouse & Meagher 1994; Smouse & al. 1999), about 0.5% is known to show female-sterile systems (Colchicaceae: Wurmbea dioica, Barrett 1992; Colchicum stevenii, Dafni & Shmida 2002; Melanthiaceae: Veratrum nigrum, Liao & al. -

Fritillaria Persica L
Turk J Bot 34 (2010) 435-440 © TÜBİTAK Research Article doi:10.3906/bot-1001-2 Male individuals in cultivated Fritillaria persica L. (Liliaceae): real androdioecy or gender disphasy? Elisa MANCUSO, Lorenzo PERUZZI* Dipartimento di Biologia dell'Università di Pisa, ITALY Received: 02.01.2010 Accepted: 28.04.2010 Abstract: In the last twenty years the growing number of studies about the reproductive biology in angiosperms has brought to light new cases of andromonoecy and androdioecy, the rarest sexual models among flowering plants. Female- sterile sexual systems, especially within order Liliales and family Liliaceae, often seem to occur in the particular form of size/age dependent sex allocation, known as “gender disphasy”. The presence of male individuals ofFritillaria persica (Liliaceae) is here documented. Comparative morphological and functional sexual expression of this species, among males and hermaphrodites, was investigated by means of flowers counting, morphometric measurements of plants and pollen-grains, pollen viability and germinability tests, and crossing experiments. The results show that hermaphrodite plants are significantly bigger and produce a higher number of flowers than males. On the other hand, there is no difference either in terms of pollen size or potential male fitness, between the 2 sex types. This suggests the occurrence of gender disphasy in this species, even if our preliminary crossing experiments seem to show an effective higher fitness of male individuals in fertilization. F. persica resulted also partially self-compatible. Our results are discussed in relation to recent findings about andromonoecious and androdioecious breeding systems within Liliales. Key words: Breeding systems, fitness, Liliales, self-compatibility Introduction androdioecy may be explained by a negative The adoption of subdioecious and submonoecious condition, in terms of genetic transmission from male sexual models is rare among angiosperms. -
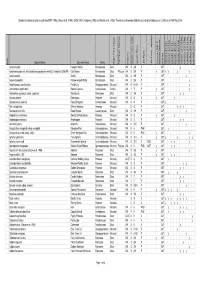
Bush Forever Species List
Species lists based on plot records from DEP (1996), Gibson et al. (1994), Griffin (1993), Keighery (1996) and Weston et al. (1992). Taxonomy and species attributes according to Keighery et al. (2006) as of 16th May 2005. Species Name Common Name Family Wd? Major Plant Group Significant Species Endemic Growth Form Code Growth Form Life Form Life Form - aquatics Common SSCP Wetland Species BFS No BRIX01 (FCT8) BRIX02 (FCT3a) BRIX03 (FCT8) BRIX04 (FCT8) BRIX05 (FCT3a) YULE01 (FCT23a) YULE02 (FCT23a) YULE03 (FCT21c) YULE04 (FCT10a) YULE05 (FCT7) Acacia huegelii Huegel's Wattle Mimosaceae Dicot WA 3 SH P 387 y Acacia lasiocarpa var. bracteolata long peduncle variant(G.J.Keighery 5026) PN Clay Moses Mimosaceae Dicot P1/p,s,e WA 3 SH P y 387 y y y y Acacia sessilis Wattle Mimosaceae Dicot WA 3 SH P 387 y Acacia stenoptera Narrow-winged Wattle Mimosaceae Dicot WA 3 SH P 387 y Acanthocarpus canaliculatus Prickle Lily Dasypogonaceae Monocot WA 4 H-SH P 387 y Actinostrobus pyramidalis Swamp Cypress Cupressaceae Conifer WA 1 T P y 387 y Adenanthos cygnorum subsp. cygnorum Woollybush Proteaceae Dicot WA 3 SH P 387 y y Agrostis plebeia Blowngrass Poaceae Monocot WA 5 G A y 387 y Agrostocrinum scabrum False Blindgrass Anthericaceae Monocot WA 4 H P 387 y * Aira caryophyllea Silvery Hairgrass Poaceae Monocot 5 G A 387 y y y Allocasuarina humilis Dwarf Sheoak Casuarinaceae Dicot WA 3 SH P 387 y Amphibromus nervosus Swamp Wallaby Grass Poaceae Monocot WA 5 G P y 387 y Amphipogon turbinatus Amphipogon Poaceae Monocot WA 5 G P 387 y y Anarthria gracilis Anarthria Restionaceae Monocot WA 6 S-R P 387 y Anigozanthos manglesii subsp. -

The Evolution of Mating Strategies in Flowering Plants Spencer C.H
trends in plant science reviews The evolution of mating strategies in flowering plants Spencer C.H. Barrett Mating strategies in flowering plants are governed by several classes of floral adaptations. Floral design and display primarily influence the quantity and quality of pollen dispersed during pollination, whereas physiological mechanisms acting in the pistil screen pollen receipt by rejecting certain male gametophytes, especially self pollen. Selective mechanisms that influence the evolution of plant mating strategies include inbreeding depression, pollen dis- counting, and the optimal allocation of resources to female and male function. New insights on the origin, evolution and function of reproductive adaptations have come from integrating studies that link microevolutionary processes to macroevolutionary patterns. he structural variation in flowers and inflorescences and reproductive mode and allows for direct testing of these models different mechanisms for deployment of male and female on the basis of the ecological and life-history associations that ac- Tgametes is remarkable when one considers that their sole company these changes. The following review examines recent pro- function is to promote reproductive success. Analysis of any plant gress in studies on the evolution of mating strategies in flowering community reveals a variety of pollination and mating systems plants and provides a discussion of the concepts and terminology that coexist under apparently similar ecological conditions. Close of the field (Box 1). relatives can reproduce in different ways just as unrelated taxa often share similar floral adaptations. How can biologists hope to Distinctive features of mating in flowering plants make sense of this variation and the evolutionary processes re- The character of mating in flowering plants is governed by dis- sponsible for the impressive reproductive versatility of flowering tinctive features of the biology and ecology of the group. -

Plants of Western Australian Granite Outcrops
JournalJournal of theof the Royal Royal Society Society of Westernof Western Australia, Australia, 80(3), 80:141-158, September 1997 1997 Plants of Western Australian granite outcrops S D Hopper1, A P Brown2 & N G Marchant3 1Kings Park and Botanic Garden, West Perth WA 6005 email: [email protected] 2Western Australian Threatened Species & Communities Unit, Department of Conservation and Land Management, PO Box 51, Wanneroo WA 6065 email: [email protected] 3Western Australian Herbarium, Department of Conservation and Land Management, PO Box 104, Como WA 6152 Abstract Outcropping granite rocks in Western Australia span a considerable climatic range, from the mediterranean south-west to inland desert and northern arid subtropics and tropics. At least 1320, and possibly 2000, plant taxa occur on Western Australian granite outcrops. Outcrop plant life is most diverse in the South West Botanical Province, with individual outcrops having up to 200 species, including many endemics not found in surrounding habitats. Species richness and local endemism declines with increasing aridity, to the point where Kimberley and Pilbara outcrops show little discontinuity in species from the surrounding landscape matrix. Outcrops are dominated by woody and herbaceous perennials, especially of the Myrtaceae, Orchidaceae, and Mimosaceae, and have an unusually rich diversity of annuals (Asteraceae, Stylidiaceae, Poaceae, Amaranthaceae etc.) compared with the flora as a whole. An unusual life form is found in resurrection plants capable of extreme desiccation and rehydration (e.g. Borya, Cheilanthes). Among woody perennials, bird pollination is frequent, and some outcrops harbour a high proportion of obligate seeder species due to the refuge from fire provided by bare rock barriers.