Revision Der Untergattung Protorina WEISE Der Gattung Oreina
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Practical Experiences in Invasive Alien Plant Control
ROSALIA Handbooks ROSALIA Handbooks Practical Experiences in Invasive Alien Plant Control Second, revised and expanded edition Invasive plant species pose major agricultural, silvicultural, human health and ecological problems worldwide, and are considered the most signifi cant threat for nature conservation. Species invading natural areas in Hungary have been described by a number of books published in the Practical Experiences in Invasive Alien Plant Control last few years. A great amount of experience has been gathered about the control of these species in some areas, which we can read about in an increasing number of articles; however, no book has been published with regards to the whole country. Invasions affecting larger areas require high energy and cost input, and the effectiveness and successfulness of control can be infl uenced by a number of factors. The development of effective, widely applicable control and eradication technologies is preceded by experiments and examinations which are based on a lot of practical experience and often loaded with negative experiences. National park directorates, forest and agricultural managers and NGOs in many parts of Hungary are combatting the spread of invasive species; however, the exchange of information and conclusion of experiences among the managing bodies is indispensable. The aim of the present volume is to facilitate this by summarizing experiences and the methods applied in practice; which, we hope, will enable us to successfully stop the further spread of invasive plant species and effectively protect our natural values. Magyarország-Szlovákia Partnerséget építünk Határon Átnyúló Együttműködési Program 2007-2013 Duna-Ipoly National Park Directorate rrosaliaosalia kkezikonyvezikonyv 3 aangng jjav.inddav.indd 1 22017.12.15.017.12.15. -

Doronicum Orientale POTW
Of interest this week at Beal... Caucasian Leopardbane Doronicum orientale Family: the Sunflower family, Asteraceae Also called Leopards Bane, Leopards Bane Daisy, W. J. Beal False Leopards Bane Botanical Garden If you are having trouble with too many leopards roaming your yard, Caucasian Leopardbane could be what you have been looking for. Its name points to the reputation the plant acquired long ago that it could repel various wild animals. It is even thought to be unappealing to cottontail rabbits. Most who plant this yellow- flowered perennial keep it for its pure yellow mid-spring color. The most vigorous specimens can attain about a meter (3 ft) in height and there are several horticultural varieties. Ours may be seen in bed 40, of the systematic collection. Caucasian Leopardbane is one of some 26 species in the genus Doronicum that are distributed across Europe, Asia, and north Africa. Doronicum orientale is found throughout southeastern Europe, across the Carpathian mountains, and in Italy. All known Doronicum species are yellow-flowered perennials that form vegetative colonies via rhizomes. Their flower heads are typical of the composite habit seen frequently throughout the sunflower family, the Asteraceae (see next page). The tendency to flower in April or May, then to disappear before the first week of July, makes Doronicum almost qualify as a spring ephemeral. In the perennial flower garden, Doronicum likes moderately moist partially shaded sites, but is tolerant of full sun. In southern localities, it depends on supplemental watering to prevent its burning away into dormancy during the hottest months of summer. Tazlaciu and Erol (1998) report that Leopards Bane appears in several medieval herbals including its listing as a diuretic, from the above-ground parts, in Turkey. -
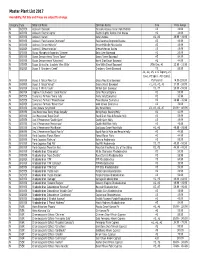
Master Plant List 2017.Xlsx
Master Plant List 2017 Availability, Pot Size and Prices are subject to change. Category Type Botanical Name Common Name Size Price Range N BREVER Azalea X 'Cascade' Cascade Azalea (Glenn Dale Hybrid) #3 49.99 N BREVER Azalea X 'Electric Lights' Electric Lights Double Pink Azalea #2 44.99 N BREVER Azalea X 'Karen' Karen Azalea #2, #3 39.99 - 49.99 N BREVER Azalea X 'Poukhanense Improved' Poukhanense Improved Azalea #3 49.99 N BREVER Azalea X 'Renee Michelle' Renee Michelle Pink Azalea #3 49.99 N BREVER Azalea X 'Stewartstonian' Stewartstonian Azalea #3 49.99 N BREVER Buxus Microphylla Japonica "Gregem' Baby Gem Boxwood #2 29.99 N BREVER Buxus Sempervirens 'Green Tower' Green Tower Boxwood #5 64.99 N BREVER Buxus Sempervirens 'Katerberg' North Star Dwarf Boxwood #2 44.99 N BREVER Buxus Sinica Var. Insularis 'Wee Willie' Wee Willie Dwarf Boxwood Little One, #1 13.99 - 21.99 N BREVER Buxus X 'Cranberry Creek' Cranberry Creek Boxwood #3 89.99 #1, #2, #5, #15 Topiary, #5 Cone, #5 Spiral, #10 Spiral, N BREVER Buxus X 'Green Mountain' Green Mountain Boxwood #5 Pyramid 14.99-299.99 N BREVER Buxus X 'Green Velvet' Green Velvet Boxwood #1, #2, #3, #5 17.99 - 59.99 N BREVER Buxus X 'Winter Gem' Winter Gem Boxwood #5, #7 59.99 - 99.99 N BREVER Daphne X Burkwoodii 'Carol Mackie' Carol Mackie Daphne #2 59.99 N BREVER Euonymus Fortunei 'Ivory Jade' Ivory Jade Euonymus #2 35.99 N BREVER Euonymus Fortunei 'Moonshadow' Moonshadow Euonymus #2 29.99 - 35.99 N BREVER Euonymus Fortunei 'Rosemrtwo' Gold Splash Euonymus #2 39.99 N BREVER Ilex Crenata 'Sky Pencil' -

Type of Presentation: Poster LS-8-P-1836 Structure And
Type of presentation: Poster LS-8-P-1836 Structure and functions of the glandular trichomes of the vegetative and reproductive organs in some Asteraceae species Muravnik L. E.1, Kostina O. V.1 1Laboratory of Plant Anatomy and Morphology, Komarov Botanical Institute of Russian Academy of Sciences, St. Petersburg, Russia Email of the presenting author: [email protected] The present work is devoted to morphological, histochemical and ultrastructural research of the trichomes covering a leaf, petiole, phyllary, and peduncle in six Senecioneae species (Doronicum macrophyllum, D. orientale, Ligularia dentata, Senecio integrifolius, S. viscosus и Tussilago farfara). The secretory structures of each species have specific morphological features. In D. macrophyllum and D. orientale a head contains one layer of the secretory cells (from two to five cells) and the long stalk (Fig. 1). Trichomes of Ligularia dentata are mainly 1-seriate, 2-seriate ones are found rarely. In S. viscosus 2–4 secretory cells form two layers of the head (Fig. 2). Leaf trichomes differ by the stalk height (from 3 to 7 cells). In S. integrifolius the trichomes of a phyllary are smaller; they have one-celled head and a short stalk. In T. farfara the trichome of the leaf and peduncle consist of a head on a long 2-seriate stalk. 3–5 layers of the secretory cells are in the head and 10–12 layers are in the stalk. All studied trichomes are capable to auto fluoresce in UV radiation without application of the markers. Yellow colour is typical for the walls of the head cells, whereas the red auto fluorescence is found in the chloroplasts of the stalk cells. -

Practical Experiences in Invasive Alien Plant Control. Rosalia Handbooks
ROSALIA Handbooks ROSALIA Handbooks Practical Experiences in Invasive Alien Plant Control Invasive plant species pose major agricultural, silvicultural, human health and ecological problems worldwide, and are considered the most signifi cant threat for nature conservation. Species invading natural areas in Hungary have been described by a number of books published in the Practical Experiences in Invasive Alien Plant Control last few years. A great amount of experience has been gathered about the control of these species in some areas, which we can read about in an increasing number of articles; however, no book has been published with regards to the whole country. Invasions affecting larger areas require high energy and cost input, and the effectiveness and successfulness of control can be infl uenced by a number of factors. The development of effective, widely applicable control and eradication technologies is preceded by experiments and examinations which are based on a lot of practical experience and often loaded with negative experiences. National park directorates, forest and agricultural managers and NGOs in many parts of Hungary are combatting the spread of invasive species; however, the exchange of information and conclusion of experiences among the managing bodies is indispensable. The aim of the present volume is to facilitate this by summarizing experiences and the methods applied in practice; which, we hope, will enable us to successfully stop the further spread of invasive plant species and effectively protect our natural values. Hungary-Slovakia Cross-border Co-operation Programme 2007-2013 Duna-Ipoly National Park Directorate Financial support for this manual has been provided by “Unified protection against invasive alien plants in sand and floodplain habitats” project. -

Reliable DNA Ploidy Determination in Dehydrated Tissues of Vascular Plants by DAPI Flow Cytometry—New Prospects for Plant Research
q 2006 International Society for Analytical Cytology Cytometry Part A 69A:273–280 (2006) Reliable DNA Ploidy Determination in Dehydrated Tissues of Vascular Plants by DAPI Flow Cytometry—New Prospects for Plant Research Jan Suda,1,2* and Pavel Travn ıcˇek1,2 1Department of Botany, Charles University, Benatska 2, Prague, Czech Republic 2Institute of Botany, Academy of Sciences, Pru˚honice 1, Czech Republic Received 30 March 2005; Revision Received 11 October 2005; Accepted 8 December 2005 Background: Only fresh plant material is generally used highly comparable with that for fresh material, allowing for rapid DNA ploidy estimation by flow cytometry (FCM). reliable DNA ploidy estimation. Deep-freezer preservation This requirement, however, substantially limits conven- substantially extended Vaccinium samples lifetime (at ient FCM application in plant biosystematics, population least to 3 years) and maintained high histogram resolu- biology, and ecology. As desiccation is a routine way for tion. sample preservation in field botany, potential utilization of Conclusions: The introduced approach eliminates the dehydrated tissues of vascular plants in FCM research was need for fresh material in many vascular plants and thus examined. opens new prospects for plant FCM. Convenient cytotype Methods: Standard DAPI protocol was employed to eval- investigation in field research and retrospective ploidy uate the performance of 60 air-dried species, spanning determination in already herbarized samples are among more than 100-fold range of nuclear DNA amounts. Multi- the principal advantages. q 2006 International Society for ploid Vaccinium subg. Oxycoccus was selected as model Analytical Cytology taxon for detailed investigation and cytotype comparison. Results: A majority of analyzed plants yielded distinct peaks with reasonable coefficients of variation after sev- Key terms: DAPI; desiccated material; DNA ploidy; flow eral months of storage at room temperature. -

Alien Vascular Plants in Iceland: Diversity, Spatial Patterns, Temporal
Flora 208 (2013) 648–673 Contents lists available at ScienceDirect Flora j ournal homepage: www.elsevier.com/locate/flora Alien vascular plants in Iceland: Diversity, spatial patterns, temporal trends, and the impact of climate change a,∗ b a Pawel Wasowicz , Ewa Maria Przedpelska-Wasowicz , Hörður Kristinsson a The Icelandic Institute of Natural History, Borgum við Norðurslóð, PO Box 180, IS-600 Akureyri, Iceland b Faculty of Biology, University of Warsaw, Miecznikowa 1, 02-096 Warszawa, Poland a r t a b i s c l e i n f o t r a c t Article history: The present study provides first comprehensive and up-to-date results on alien plant taxa in Iceland Received 24 May 2013 since 1967. We evidenced the presence of 336 alien vascular plant taxa, including 277 casuals and 59 Accepted 26 September 2013 naturalised taxa, two being invasive. The distribution of the alien flora exhibits a clear spatial pattern Available online 6 October 2013 showing hotspots of occurrence and diversity within areas of major settlement centres. Altitude above sea level and temperature-related variables proved to be the most important factors shaping alien plant Keywords: distribution in Iceland. Predictive modelling evidenced that arctic areas of Iceland and the Central High- Alien flora lands are under serious risk of alien plant invasion due to climate change. The results provide crucial Iceland information for alien and invasive plant management and contribute data for meta-analyses of invasion Climate change processes worldwide. Invasive species Predictive modelling © 2013 Elsevier GmbH. All rights reserved. Maxent Introduction decades northern plant taxa, well adapted to cold environments will come under pressure and have to withdraw from their natural Only a small percent of alien plant taxa have a potential to environments. -

Volatile Constituents of the Flowers and Leaves-Stems of Three Doronicum Taxa from Turkey
Asian Journal of Chemistry Vol. 21, No. 2 (2009), 1225-1229 Volatile Constituents of the Flowers and Leaves-Stems of three Doronicum taxa from Turkey KURTULUS AKPINAR, NURI Y ILDIRIM, OSMAN ÜÇÜNCÜ, NURAN Y AYLI, SALIH TERZIOGLU† and NURETTIN Y AYLI* Department of Chemistry, Faculty of Arts and Sciences Karadeniz Technical University, 61080 Trabzon, Turkey Fax: (90)(462) 3253196; Tel: (90)(462)3772486 E-mail: [email protected] The composition of the essential oils obtained from fresh flowers and leaves-stems of 3 Doronicum taxa, 2 of them are endemic to Turkey, have been studied. The essential oils were separately obtained from hydro- distillation in a clevenger-type apparatus and their analyses were perfor- med by GC and GC-MS. A range of 14-27 compounds were identified, constituting over 89.3 to 97.7 % of total oil composition. Sesquiterpene hydrocarbons were shown to be the main group of constituents of all 3 Doronicum taxa. The main component of the flowers and leaves-stems in the essential oils of Doronicum orientale and Doronicum bithynicum ssp. sparsipilosum was (E)-β-farnesene (41.1, 35.7, 47.5 and 55.4 %, respectively); while (E)-caryophyllene (24.2 and 52.7 %) was the major compound in the essential oils of Doronicum macrolepis. Key Words: Doronicum orientale, Doronicum bithynicum subsp. sparsipilosum, Doronicum macrolepis, Essential oil, GC, GC-MS. INTRODUCTION Doronicum L. (Asteraceae) represented with 15 native taxa of which 14 are in species level in Turkey1. They are perennial herbs with short usually horizontal rhizome and 8 of the taxa are endemic to Turkey1. -

NEW LOCALITIES of the SPECIES Doronicum Orientale HOFFM
View metadata, citation and similar papers at core.ac.uk brought to you by CORE NAT. CROAT. VOL. 8 No 2 87¿93 ZAGREB June 30, 1999 ISSN 1330-0520 UDK 582.998:581.93(497.5) NEW LOCALITIES OF THE SPECIES Doronicum orientale HOFFM. (Asteraceae) IN CROATIA MIRKO TOMA[EVI]1, IVICA SAMARD@I]2 & LJUDEVIT ILIJANI]3 1V. Ma~eka 9, 34000 Po`ega, Croatia 2»Po`ega« Forestry Department, 34000 Po`ega, Croatia 3Institute of Botany, Dept. of Biology, Faculty of Science, Maruli}ev trg 20/II, 10000 Zagreb, Croatia Toma{evi}, M., Samard`i}, I. & Ilijani}, Lj.: New localities of the species Doronicum orientale Hoffm. (Asteraceae) in Croatia, Nat. Croat., Vol. 8, No. 2., 87–93, 1999, Zagreb. The species Doronicum orientale Hoffm. (= D. caucasium Bieb.) is a rare plant in the flora of Croa- tia, and has been protected by law since 1958. Hitherto, two localities were known: on Medvednica Mountain near Zagreb (UTM WL 78), and on Psunj Mountain in Slavonia (UTM XL 93). This re- search reports its presence also on Papuk Mountain (UTM YL 03) in a thermophilous white oak forest and on Po`e{ka Gora Mountain (UTM YL 01) in a sessile oak and hornbeam forest. Its distri- bution in Croatia is shown on the map (Fig. 1). Key words: Doronicum orientale, distribution, Croatia Toma{evi}, M., Samard`i}, I. & Ilijani}, Lj: Nova nalazi{ta vrste Doronicum orientale Hoffm. (Asteraceae) u Hrvatskoj, Nat. Croat., Vol. 8, No. 2., 87–93, 1999, Zagreb. Vrsta Doronicum orientale Hoffm. (= D. caucasicum Bieb.) rijetka je biljka hrvatske flore, zakonom za{ti}ena od 1958. -

Adatok a Hazai Dráva Menti Síkság Flórájához
105 KITAIBELIA XVIII. évf. 1-2. szám pp.: 105–124. Debrecen 2013 Adatok a hazai Dráva menti síkság flórájához 1,2 KEVEY BALÁZS (1) Pécsi Tudományegyetem, (1) Növényrendszertani és Geobotanikai Tanszék; (2) Szőlészeti és Agrobotanikai Tanszék; 7624 Pécs, Ifjúság u. 6., [email protected] Bevezetés A hazai Dráva-ártér növényvilágáról először BOROS (1924) közölt élethű képet. HORVÁT (1942) flóraműve a Dráva-sík baranyai szakaszára is kiterjed, amelyhez a szerző több alkalommal is közöl pótlásokat (HORVÁT 1972, 1972-1973, 1974, 1976 stb.). A szigorú határőrizet miatt évtizedeken át alig lehetett megközelíteni az országhatárhoz közel eső részeket, így azok növényvilágáról sokáig csak hiányos ismeretekkel rendelkeztünk. Mindezek ellenére BORHIDI (1958a, 1958b) és HÉJJAS – BORHIDI (1960) a Dráva somogyi, VÖRÖSS (1964a, 1964b, 1965, 1968, 1971, 1974) pedig a baranyai szakaszáról közöl értékes florisztikai és cönológiai adatokat. Magam 1973-ban kezdtem kutatni a Dráva-sík növényvilágát, elsősorban az erdőket. A cönológiai felvételek készítése közben számos értékes florisztikai adatra bukkantam, amelyek egy részét korábban már közöltem (vö. KEVEY in PRISZTER 1980; KEVEY 1980, 1983, 1985, 1987-1988, 1989, 1993, 1995, 2001a, 2001b, 2001c, 2004a, 2004b; KEVEY in DÉNES et al. 1996–1997; KEVEY – KIRÁLY 2002; KEVEY – BARTHA 2010a, 2010b, 2010c;). Az utóbbi két évtizedben több kutató is végzett felméréseket a Dráva-síkon (DÉNES 1996, 1997, 1998; DÉNES et al. 1996-1997, 2008; DÉNES – ORTMANN-AJKAI 1998, 2006; ORTMANN-AJKAI 1998a, 1998b; LÁJER 1999; CSIKY et al. 2004, 2005, 2006; JUHÁSZ – DÉNES 2005, 2006; CSIKY – OLÁH 2006; PÁL et al. 2006, 2010; PÁL – PINKE 2007; BÁTORI et al. 2010; LENGYEL et al. 2012). Számos florisztikai adat olvasható ki a korábban közölt cönológiai táblázataimból is (vö. -

Reproduction and Identification of Root-Knot Nematodes on Perennial Ornamental Plants in Florida
REPRODUCTION AND IDENTIFICATION OF ROOT-KNOT NEMATODES ON PERENNIAL ORNAMENTAL PLANTS IN FLORIDA By ROI LEVIN A THESIS PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE UNIVERSITY OF FLORIDA 2005 Copyright 2005 by Roi Levin ACKNOWLEDGMENTS I would like to thank my chair, Dr. W. T. Crow, and my committee members, Dr. J. A. Brito, Dr. R. K. Schoellhorn, and Dr. A. F. Wysocki, for their guidance and support of this work. I am honored to have worked under their supervision and commend them for their efforts and contributions to their respective fields. I would also like to thank my parents. Through my childhood and adult years, they have continuously encouraged me to pursue my interests and dreams, and, under their guidance, gave me the freedom to steer opportunities, curiosities, and decisions as I saw fit. Most of all, I would like to thank my fiancée, Melissa A. Weichert. Over the past few years, she has supported, encouraged, and loved me, through good times and bad. I will always remember her dedication, patience, and sacrifice while I was working on this study. I would not be the person I am today without our relationship and love. iii TABLE OF CONTENTS page ACKNOWLEDGMENTS ................................................................................................. iii LIST OF TABLES............................................................................................................. vi LIST OF FIGURES .......................................................................................................... -

THE VASCULAR PLANTS of BRITISH COLUMBIA Part 1
TABLE OF CONTENTS ACKNOWLEDGEMENTS.................................................................... iv INTRODUCTION ........................................................................... 1 Format................................................................................ 1 Data Base ............................................................................. 3 Taxonomic Concepts .................................................................... 3 GYMNOSPERMS ........................................................................... 4 DICOTYLEDONS (ACERACEAE THROUGH CUCURBITACEAE) ................................ 9 REFERENCES............................................................................. 165 EXCLUDED SPECIES ...................................................................... 181 INDEX .................................................................................... 188 SEQ 5601 JOB FLORA4-000-009 PAGE-0001 CAMPANULACEAE REVISED 19JUL00 AT 15:32 BY BC DEPTH: 65 PICAS WIDTH 44 PICAS COLOR LEVEL 1 Campanulaceae CAMPANULA 1. Plants usually over 5 dm tall; stems leafy; flowers in a terminal cluster; introduced species. 2. Plants biennials, hispid-hairy; calyx with a conspicuous appendage at each sinus; corollas large, usually 3.5-5 cm long ......................................................... C. medium 2. Plants perennials, glabrous or sparsely pubescent; calyx without appendages; corollas smaller, usually 2-3.5 cm long. 3. Leaves linear to linear-lanceolate, finely crenate, the lower stem leaves subsessile