Tree Diversity in Sub-Montane and Lower Montane Primary Rain Forests in Central Sulawesi
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Take Another Look
Take Contact Details Another SUNSHINE COAST REGIONAL COUNCIL Caloundra Customer Service Look..... 1 Omrah Avenue, Caloundra FRONT p: 07 5420 8200 e: [email protected] Maroochydore Customer Service 11-13 Ocean Street, Maroochydore p: 07 5475 8501 e: [email protected] Nambour Customer Service Cnr Currie & Bury Street, Nambour p: 07 5475 8501 e: [email protected] Tewantin Customer Service 9 Pelican Street, Tewantin p: 07 5449 5200 e: [email protected] YOUR LOCAL CONTACT Our Locals are Beauties HINTERLAND EDITION HINTERLAND EDITION 0 Local native plant guide 2 What you grow in your garden can have major impact, Introduction 3 for better or worse, on the biodiversity of the Sunshine Native plants 4 - 41 Coast. Growing a variety of native plants on your property can help to attract a wide range of beautiful Wildlife Gardening 20 - 21 native birds and animals. Native plants provide food and Introduction Conservation Partnerships 31 shelter for wildlife, help to conserve local species and Table of Contents Table Environmental weeds 42 - 73 enable birds and animals to move through the landscape. Method of removal 43 Choosing species which flower and fruit in different Succulent plants and cacti 62 seasons, produce different types of fruit and provide Water weeds 70 - 71 roost or shelter sites for birds, frogs and lizards can greatly increase your garden’s real estate value for native References and further reading 74 fauna. You and your family will benefit from the natural pest control, life and colour that these residents and PLANT TYPE ENVIRONMENTAL BENEFITS visitors provide – free of charge! Habitat for native frogs Tall Palm/Treefern Local native plants also improve our quality of life in Attracts native insects other ways. -

IFE and Wlldll ITAT of AMERICAN SA VIRONMENT and ECOLO
IFE AND WlLDLl ITAT OF AMERICAN SA VIRONMENT AND ECOLO By A. Binion Amerson, Jr., W. Arthur Whistler, and Terry D. Schwaner Environment Consultants, Inc., Dallas, Texas Edited by Richard C. Banks U.S. Fish and Wildlife Service Washington, D.C. UNITED STATES DEPARTMENT OF T E INTERIOR FISH AND WILDLIFE SERVICE Washington, D.C. e 1982 Foreword A survey of the status of the wildlife and wildlife habitat of American Samoa. an unincorporated Territory of the United States. was recommended by administrative officials of the U.S. Fish and Wildlife Service (FWS) in the early 1970s .Environ- ment Consultants. Inc . (ECI). based in Dallas. Texas. was selected to conduct a 2-year survey with A .Binion Amerson. Jr., as Principal Investigator . The contract was administered through the Division of Federal Aid in FWS Region I. Portland. Oregon . The primary objectives of the survey were (1) to define the major ecosystems and to inventory their physical components. vegetation. and wildlife constituents; (2) to prepare maps of these ecosystems; (3) to identify any threatened or endangered species of wildlife; and (4) to recommend wildlife management opportunities and needs . The report of the survey was to be in two parts . The first was to be a non-technical account suitable for wide general distribution; the second was to include the technical aspects of the data and data gathering. with accounts of the wildlife species . This volume represents the first part of ECI's report . The final report submitted by ECI contained more than 1. 200 pages. 200 figures. and 110 tables. many of thelatter several pages long . -

Endiandric Acid Derivatives and Other Constituents of Plants from the Genera Beilschmiedia and Endiandra (Lauraceae)
Biomolecules 2015, 5, 910-942; doi:10.3390/biom5020910 OPEN ACCESS biomolecules ISSN 2218-273X www.mdpi.com/journal/biomolecules/ Review Endiandric Acid Derivatives and Other Constituents of Plants from the Genera Beilschmiedia and Endiandra (Lauraceae) Bruno Ndjakou Lenta 1,2,*, Jean Rodolphe Chouna 3, Pepin Alango Nkeng-Efouet 3 and Norbert Sewald 2 1 Department of Chemistry, Higher Teacher Training College, University of Yaoundé 1, P.O. Box 47, Yaoundé, Cameroon 2 Organic and Bioorganic Chemistry, Chemistry Department, Bielefeld University, P.O. Box 100131, 33501 Bielefeld, Germany; E-Mail: [email protected] 3 Department of Chemistry, University of Dschang, P.O. Box 67, Dschang, Cameroon; E-Mails:[email protected] (J.R.C.); [email protected] (P.A.N.-E.) * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +2376-7509-7561. Academic Editor: Jürg Bähler Received: 3 March 2015 / Accepted: 6 May 2015 / Published: 14 May 2015 Abstract: Plants of the Lauraceae family are widely used in traditional medicine and are sources of various classes of secondary metabolites. Two genera of this family, Beilschmiedia and Endiandra, have been the subject of numerous investigations over the past decades because of their application in traditional medicine. They are the only source of bioactive endiandric acid derivatives. Noteworthy is that their biosynthesis contains two consecutive non-enzymatic electrocyclic reactions. Several interesting biological activities for this specific class of secondary metabolites and other constituents of the two genera have been reported, including antimicrobial, enzymes inhibitory and cytotoxic properties. This review compiles information on the structures of the compounds described between January 1960 and March 2015, their biological activities and information on endiandric acid biosynthesis, with 104 references being cited. -
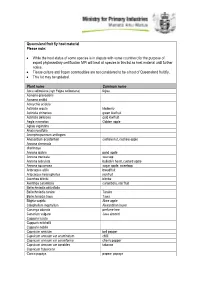
MPI Queensland Fruit Fly Host Material List
Queensland fruit fly host material Please note: While the host status of some species is in dispute with some countries; for the purpose of export phytosanitary certification MPI will treat all species in this list as host material until further notice. Tissue culture and frozen commodities are not considered to be a host of Queensland fruit fly. This list may be updated. Plant name Common name Acca sellowiana (syn Feijoa sellowiana) feijoa Acmena graveolens Acmena smithii Acroychia acidula Actinidia arguta kiwiberry Actinidia chinensis green kiwifruit Actinidia deliciosa gold kiwifruit Aegle marmelos Golden apple Aglaia sapindina Alyxia ruscifolia Amorphospermum antilogum Anacardium occidentale cashew nut, cashew apple Annona cherimola cherimoya Annona glabra pond apple Annona muricata soursop Annona reticulata bullock's heart, custard apple Annona squamosa sugar apple, sweetsop Artocarpus altilis breadfruit Artocarpus heterophyllus jackfruit Averrhoa bilimbi blimbe Averrhoa carambola carambola, star fruit Beilschmiedia obtusifolia Beilschmiedia taraire Taraire Beilschmiedia tawa Tawa Blighia sapida Akee apple Calophyllum inophyllum Alexandrian laurel Cananga odorata perfume tree Canarium vulgare Java almond Capparis lucida Capparis mitchellii Capparis nobilis Capsicum annuum bell pepper Capsicum annuum var acuminatum chilli Capsicum annuum var cerasiforme cherry pepper Capsicum annuum var conoides tabasco Capsicum frutescens Carica papaya papaw, papaya Carica pentagona babaco Carissa ovata Casimiroa edulis white sapote Cassine australia Castanospora alphandii Chrysophyllum cainito caimito, star apple Cissus sp Citrullus lanatus Watermelon Citrus spp. Citrus aurantiifolia West indian lime Citrus aurantium Seville orange Citrus grandis (syn maxima) pummelo Citrus hystrix kaffir lime Citrus jambhiri rough lemon Citrus latifolia Tahatian lime Citrus limetta sweet lemon tree Citrus limon lemon Citrus maxima (syn grandis) pomelo, shaddock Citrus medica citron Citrus meyeri Meyer lemon Citrus reticulata mandarin Citrus reticulata var. -

Threatened Species of Wilsons and Coopers Creek
Listed below are species recorded from the project areas of Goonengerry Landcare and Wilsons Creek Huonbrook Landcare groups. Additional species are known from adjacent National Parks. E = Endangered V = Vulnerable BCA - Biodiversity Conservation Act 2016 EPBC - Environment Protection and Biodiversity Conservation Act 1999 Threatened Species of Wilsons and Coopers Creek SOS - Saving our Species Scientific name Common name TSC Act status EPBC Act status SOS stream Wilsons Creek and Coopers Creek are tributaries of the Wilsons River on the Far North Coast of New South Wales. Within the South East Queensland Bioregion, the native flora and fauna of PLANTS this region are among the most diverse in Australia. In the catchment areas of the Wilsons and Corokia whiteana Corokia V V Keep watch Coopers Creek 50 threatened species of flora and fauna can be found and 2 endangered Davidsonia johnsonii Smooth Davidson's Plum E E Site managed ecological communities. Desmodium acanthocladum Thorny Pea V V Site managed What is a threatened species? Diploglottis campbellii Small-leaved Tamarind E E Site managed Plants and animals are assessed on the threats that face them and the level to which they are at Doryanthes palmeri Giant Spear Lily V Keep watch risk of extinction. If the risk is high they are listed in legislation and conservation actions are Drynaria rigidula Basket Fern E Partnership developed for their protection. There are almost 1000 animal and plant species at risk of Elaeocarpus williamsianus Hairy Quandong E E Site managed extinction in NSW. Endiandra hayesii Rusty Rose Walnut V V Data deficient A species is considered threatened if: Endiandra muelleri subsp. -

Coolum Street Tree Strategy
Coolum Street tree strategy Description of plan area and land use Major opportunities and constraints Street tree planting strategies Coolum provides a casual seaside setting for urban residential living and The best street tree planting opportunities in the Coolum locality exist in the tourism. The plan area covers the established seaside suburbs of Coolum, form of planting nodes, where large trees or groups of trees can be readily Street tree plantings reinforce the casual beachside character of Point Arkwright, Yaroomba and Mount Coolum, the newer residential accommodated. the precinct and are considerate of existing scenic amenity and the developments of the Boardwalk and the Town of Seaside, and the Coolum Significant opportunity to plant succession trees in areas of foreshore that preservation of existing water views. Industrial Park west of the Sunshine Motorway. Covering 1880 hectares, the intersect with streets is evident. Planting trees in areas of foreshore where landscape and extent of tree cover varies considerably, yet a relaxed urban Streetscapes predominately exhibit mixed native species plantings. no water views exist is a key priority. coastal character is consistent throughout the locality. Gateways and node plantings on Coolum–Yandina Road and the Additional street tree planting opportunities include the potential to infill Esplanade enhance the sense of arrival to the precinct. Trees and landscape character major avenues with new street trees, build canopy on parkland perimeters (fronting local streets) and provide shade to numerous pedestrian routes Succession trees are planted wherever there are no water views and The Coolum plan area contains many significant natural features including and pathways. -

Severe Tropical Cyclone Larry
The effects of Severe Tropical Cyclone Larry on rainforest vegetation and understorey microclimate adjacent to powerlines, highways and streams in the Wet Tropics World Heritage Area Catherine L. Pohlman and Miriam Goosem School of Earth and Environmental Sciences, James Cook University Supported by the Australian Government’s Marine and Tropical Sciences Research Facility Project 4.9.3 Impacts of urbanisation on North Queensland environments: management and remediation © James Cook University ISBN 978-1-921359-12-5 This report should be cited as: Pohlman, C. and Goosem, M. (2007) The effects of Severe Tropical Cyclone Larry on rainforest vegetation and understorey microclimate adjacent to powerlines, highways and streams in the Wet Tropics World Heritage Area. Report to the Marine and Tropical Sciences Research Facility. Reef and Rainforest Research Centre Limited, Cairns (47pp.). Made available online by the Reef and Rainforest Research Centre on behalf of the Australian Government’s Marine and Tropical Sciences Research Facility. The Australian Government’s Marine and Tropical Sciences Research Facility (MTSRF) supports world-class, public good research. The MTSRF is a major initiative of the Australian Government, designed to ensure that Australia’s environmental challenges are addressed in an innovative, collaborative and sustainable way. The MTSRF investment is managed by the Department of the Environment, Water, Heritage and the Arts (DEWHA), and is supplemented by substantial cash and in-kind investments from research providers and interested third parties. The Reef and Rainforest Research Centre Limited (RRRC) is contracted by DEWHA to provide program management and communications services for the MTSRF. This publication is copyright. The Copyright Act 1968 permits fair dealing for study, research, information or educational purposes subject to inclusion of a sufficient acknowledgement of the source. -

A Review of Endiandric Acid Analogues
Available online on www.ijppr.com International Journal of Pharmacognosy and Phytochemical Research 2015; 7(4); 844-856 ISSN: 0975-4873 Review Article A Review of Endiandric Acid Analogues Salleh WMNHW1, Ahmad F1*, Khong HY2, Zulkifli RM3 1Department of Chemistry, Faculty of Science, Universiti Teknologi Malaysia (UTM), 81310 Skudai, Johor, Malaysia 2School of Chemistry and Environmental Studies, Faculty of Applied Sciences, Universiti Teknologi MARA (UiTM) 94300 Kota Samarahan, Sarawak, Malaysia 3Department of Biosciences and Health Sciences, Faculty of Biosciences and Medical Engineering, Universiti Teknologi Malaysia (UTM), 81310 Skudai, Johor, Malaysia. Available Online: 31st July, 2015 ABSTRACT The genus Beilschmiedia and Endiandra belonging to Lauraceae family have been used as folk medicines in treating uterine tumors, rheumatism, pulmonary diseases, dysentery and headache. This report briefly the current information and update on isolated endiandric acid analogues and their pharmacological studies. The informations on both genus were gathered via internet using scientific databases such as Google Scholar, MedlinePlus, Pubmed, SciFinder, Scopus and Web of Science. A literature survey shows that both genus are mainly known for their endiandric acid analogues with antiapoptotic proteins, which emphasize the existence of various potential leads to develop new anticancer agents. Modern pharmacology studies have also demonstrated that their phytochemicals possess wide pharmacological activities, which are anti-inflammatory, cytotoxic, antibacterial, antiplasmodial and antitubercular activities. Keywords: Endiandric acid, Beilschmiedia, Endiandra, Phytochemistry, Pharmacology INTRODUCTION throughout tropical to subtropical latitudes especially The World Health Organization (WHO) estimated that 80% Southeast Asia and tropical America. In Malaysia, it is of the populations of the developing countries rely on known as ‘medang’ or ‘tejur’ and contribute about 213 traditional medicines, mostly plant-based drugs, for their species from 16 genera7-8. -

Juglandaceae (Walnuts)
A start for archaeological Nutters: some edible nuts for archaeologists. By Dorian Q Fuller 24.10.2007 Institute of Archaeology, University College London A “nut” is an edible hard seed, which occurs as a single seed contained in a tough or fibrous pericarp or endocarp. But there are numerous kinds of “nuts” to do not behave according to this anatomical definition (see “nut-alikes” below). Only some major categories of nuts will be treated here, by taxonomic family, selected due to there ethnographic importance or archaeological visibility. Species lists below are not comprehensive but representative of the continental distribution of useful taxa. Nuts are seasonally abundant (autumn/post-monsoon) and readily storable. Some good starting points: E. A. Menninger (1977) Edible Nuts of the World. Horticultural Books, Stuart, Fl.; F. Reosengarten, Jr. (1984) The Book of Edible Nuts. Walker New York) Trapaceae (water chestnuts) Note on terminological confusion with “Chinese waterchestnuts” which are actually sedge rhizome tubers (Eleocharis dulcis) Trapa natans European water chestnut Trapa bispinosa East Asia, Neolithic China (Hemudu) Trapa bicornis Southeast Asia and South Asia Trapa japonica Japan, jomon sites Anacardiaceae Includes Piastchios, also mangos (South & Southeast Asia), cashews (South America), and numerous poisonous tropical nuts. Pistacia vera true pistachio of commerce Pistacia atlantica Euphorbiaceae This family includes castor oil plant (Ricinus communis), rubber (Hevea), cassava (Manihot esculenta), the emblic myrobalan fruit (of India & SE Asia), Phyllanthus emblica, and at least important nut groups: Aleurites spp. Candlenuts, food and candlenut oil (SE Asia, Pacific) Archaeological record: Late Pleistocene Timor, Early Holocene reports from New Guinea, New Ireland, Bismarcks; Spirit Cave, Thailand (Early Holocene) (Yen 1979; Latinis 2000) Rincinodendron rautanenii the mongongo nut, a Dobe !Kung staple (S. -

Cryptocarya Nitens (Lauraceae), a New Species Record for Singapore
Gardens’ Bulletin Singapore 67(2): 253–259. 2015 253 doi: 10.3850/S2382581215000204 Cryptocarya nitens (Lauraceae), a new species record for Singapore R.P.J. de Kok 275 Cricklewood Lane, London, NW2 2JJ, U.K. [email protected] ABSTRACT. Cryptocarya nitens (Blume) Koord. & Valeton is newly recorded for Singapore. It was discovered during surveys in Bukit Timah Nature Reserve and the Nee Soon Swamp Forest. A description is given, together with a key based mostly on vegetative characters for all Cryptocarya species occurring in Singapore. A short overview of the Lauraceae of Singapore shows that, in total, 57 species belonging to 14 genera have been recorded, of which 47 species in 13 genera are native. Cryptocarya nitens is lectotypified in addition to two of its synonyms. Keywords. Cryptocarya, Endiandra Introduction Species of the Lauraceae (Laurel Family) are of major ecological and economic importance in Southeast Asia as they comprise a major part of almost any forest in the region. The family is very complex, posing many challenging taxonomic and systematic questions, and for various reasons the identification of species is problematic. Some systematic work has been done, for instance Kostermans (1952, 1957a, 1957b, 1964) wrote a series of papers on various genera or parts of genera during his career, and he also dealt with some aspects of the higher taxonomy. The most recent overview of the family as a whole was by Rohwer (1993). In Eastern Asia and Australasia, only for Peninsular Malaysia (Kochummen, 1989), China (Li Xiwen et al., 2008) and Australia (Le Cussan & Hyland, 2007) are recent treatments available, dealing with all species occurring in their territories. -

High Performance Liquid Chromatography in Phytochemical Analysis CHROMATOGRAPHIC SCIENCE SERIES
High Performance Liquid Chromatography in Phytochemical Analysis CHROMATOGRAPHIC SCIENCE SERIES A Series of Textbooks and Reference Books Editor: JACK CAZES 1. Dynamics of Chromatography: Principles and Theory, J. Calvin Giddings 2. Gas Chromatographic Analysis of Drugs and Pesticides, Benjamin J. Gudzinowicz 3. Principles of Adsorption Chromatography: The Separation of Nonionic Organic Compounds, Lloyd R. Snyder 4. Multicomponent Chromatography: Theory of Interference, Friedrich Helfferich and Gerhard Klein 5. Quantitative Analysis by Gas Chromatography, Josef Novák 6. High-Speed Liquid Chromatography, Peter M. Rajcsanyi and Elisabeth Rajcsanyi 7. Fundamentals of Integrated GC-MS (in three parts), Benjamin J. Gudzinowicz, Michael J. Gudzinowicz, and Horace F. Martin 8. Liquid Chromatography of Polymers and Related Materials, Jack Cazes 9. GLC and HPLC Determination of Therapeutic Agents (in three parts), Part 1 edited by Kiyoshi Tsuji and Walter Morozowich, Parts 2 and 3 edited by Kiyoshi Tsuji 10. Biological/Biomedical Applications of Liquid Chromatography, edited by Gerald L. Hawk 11. Chromatography in Petroleum Analysis, edited by Klaus H. Altgelt and T. H. Gouw 12. Biological/Biomedical Applications of Liquid Chromatography II, edited by Gerald L. Hawk 13. Liquid Chromatography of Polymers and Related Materials II, edited by Jack Cazes and Xavier Delamare 14. Introduction to Analytical Gas Chromatography: History, Principles, and Practice, John A. Perry 15. Applications of Glass Capillary Gas Chromatography, edited by Walter G. Jennings 16. Steroid Analysis by HPLC: Recent Applications, edited by Marie P. Kautsky 17. Thin-Layer Chromatography: Techniques and Applications, Bernard Fried and Joseph Sherma 18. Biological/Biomedical Applications of Liquid Chromatography III, edited by Gerald L. Hawk 19. -

A Biological Assessment of the Wapoga River Area of Northwestern Irian Jaya, Indonesia
Rapid Assessment Program 14 RAP Bulletin of Biological Assessment A Biological Assessment of the Wapoga River Area of Northwestern Irian Jaya, Indonesia Andrew L. Mack and Leeanne E. Alonso, Editors CENTER FOR APPLIED BIODIVERSITY SCIENCE (CABS) CONSERVATION INTERNATIONAL INDONESIAN NATIONAL INSTITUTE OF SCIENCES (LIPI) BANDUNG TECHNOLOGY INSTITUTE (ITB) UNIVERSITY OF CENDERAWASIH (UNCEN) PERLINDUNGAN DAN KONSERVASI ALAM (PKA) BADAN PENGEMBANGAN DAN PEMBANGUNAN RAP BULLETIN OF BIOLOGICAL ASSESSMENT FOURTEEN DAERAH January (BAPPEDA) 2000 1 Rapid Assessment Program 14 RAP Bulletin of Biological Assessment A Biological Assessment of the Wapoga River Area of Northwestern Irian Jaya, Indonesia Andrew L. Mack and Leeanne E. Alonso, Editors CENTER FOR APPLIED BIODIVERSITY SCIENCE (CABS) CONSERVATION INTERNATIONAL INDONESIAN NATIONAL INSTITUTE OF SCIENCES (LIPI) BANDUNG TECHNOLOGY INSTITUTE (ITB) UNIVERSITY OF CENDERAWASIH (UNCEN) PERLINDUNGAN DAN KONSERVASI ALAM (PKA) BADAN PENGEMBANGAN DAN PEMBANGUNAN RAP BULLETIN OF BIOLOGICAL ASSESSMENT FOURTEEN DAERAH January (BAPPEDA) 2000 1 RAP Bulletin of Biological Assessment is published by: Conservation International Center for Applied Biodiversity Science Department of Conservation Biology 2501 M Street NW, Suite 200 Washington, DC 20037 USA 202-429-5660 tel 202-887-0193 fax www.conservation.org Editors: Andrew L. Mack and Leeanne E. Alonso Design: Glenda P. Fábregas Map: Dan Polhemus and Leeanne E. Alonso Cover photograph: Michael Moore Translations: Iwan Wijayanto and Suer Surjadi Conservation International